
Mantidactylus glosi, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504425 |
|
persistent identifier |
https://treatment.plazi.org/id/6A1878F4-413F-45DF-BF32-0EA7389A6452 |
|
taxon LSID |
lsid:zoobank.org:act:6A1878F4-413F-45DF-BF32-0EA7389A6452 |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus glosi |
| status |
sp. nov. |
Mantidactylus glosi sp. nov.
Identity and justification.—This species has been considered as confirmed candidate species M. sp. 24 by Vieites et al. (2009), and M. sp. Ca24 by Perl et al. (2014). It was depicted as ‘ Mantidactylus sp. aff. biporus “Ranomafana”’ by Glaw and Vences (2007) and Schmidt et al. (2009) [tadpoles]. A lineage of the M. biporus clade with strong divergence in 16S and not immediately closely related to another lineage in the phylogenomic tree (i.e. sister to a subclade with four other species). Also characterized by differentiation in Rag-1 from other species in the M. biporus clade, and therefore here seen as distinct species.
Holotype.— ZSM 434/2006 ( ZCMV 3366 ), adult male, collected by M. Vences, Y. Chiari, T. Rajofiarison, E. Rajeriarison, P. Bora, and T. Razafindrabe on 26 February 2006 at Ambatovory (upstream river from Ambatolahy), Ranomafana National Park (21.23798°S, 047.42478°E, 966 m a.s.l.), Vatovavy-Fitovinany Region, Madagascar. A 16S barcode sequence of the holotype is available from GenBank (accession FJ559255 View Materials ). GoogleMaps
Paratypes.—A total of four paratypes: ZSM 433/2006 ( ZCMV 3364 ), adult female, with same collection data as holotype GoogleMaps ; UADBA uncatalogued ( ZCMV 3365 , ZCMV 3367 ), two specimens of unknown sex and maturity, with same collection data as the holotype GoogleMaps ; UADBA 20667 ( FGMV 2002.254 ), unknown sex and maturity, collected by F. Glaw, M. Puente, L. Raharivololoniaina, M. Thomas, and D.R. Vieites on 16 January 2003 in Ranomafana National Park.
Diagnosis.— Mantidactylus glosi sp. nov. is a member of the M. biporus clade and is sister to the assemblage of all other species in the clade according to our phylogenomic analysis. See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of small body size (male SVL 21 mm, female SVL 25 mm), slightly granular dorsal skin with weakly expressed and sometimes discontinuous but clearly recognisable dorsolateral ridges, large tympanum size in males (13% of SVL), presence of white spots on flanks in at least some individuals, and absence of a white marking on the snout tip, distinguishes M. glosi sp. nov. from species of the M. betsileanus , M. curtus , M. fergusoni , M. tricinctus , and M. ulcerosus clades. M. inaudax ( M. inaudax clade) differs by larger body size, smoother dorsal skin and longer hindlimbs; M. biporus differs by larger body size and smoother dorsal skin; M. augustini differs by longer hindlimbs; M. bletzae differs by more developed foot webbing and longer hindlimbs; M. brevirostris has a less developed foot webbing, smoother dorsal skin, and longer hindlimbs; M. eulenbergeri has a smoother dorsal skin ( Table 4 View TABLE 4 ). For a distinction from other new species in the M. stelliger and M. inaudax clades, see the diagnoses in the respective species accounts below.
A full list of molecular diagnostic sites in the 16S gene of M. glosi sp. nov. in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Description of the holotype.—Adult male in good state of preservation ( Fig. 62 View FIGURE 62 ). Part of right thigh muscle removed as tissue sample. Body rather stout. Head slightly wider than body. Snout rounded to weakly pointed in dorsal view. Nostrils directed laterally, not protuberant.
Nostrils nearer to tip of the snout than to eye. Canthus rostralis not recognisable. Loreal region concave.
Tympanum distinct, large, rounded, its horizontal diameter about 88% of eye diameter. Supratympanic fold present, beginning straight, with a distinct almost 90° bend midway towards jaw / forelimb insertion. Tongue ovoid, distinctly bifid. Maxillary teeth present. Vomerine teeth present in two rounded aggregations, positioned posterolateral to choanae. Choanae rounded. Subarticular tubercles single. Inner and outer metacarpal tubercles present. Fingers without webbing. Relative length of fingers: I<II<IV<III. Finger discs very slightly enlarged. Nuptial pads absent. Foot of similar length as tibia (102%). Lateral metatarsalia separated. Inner and outer metatarsal tubercles present. Webbing formula: 1(1), 2i(1.5), 2e(1), 3i(2), 3e(2), 4i(2.5), 4e(2), 5(1). Relative length of toes: I<II<V<III<IV. Skin on the upper surface quite smooth in preservative with a few granules on the lower flank; in life, as in photographed specimens of this species, likely with small scattered granules and one pair of poorly developed continuous dorsolateral folds—these are weakly recognisable in preservative. Ventral side smooth. Femoral glands present, made up by a distinct but relatively small distal ulcerous macrogland internally consisting of five large granules. Proximal granular gland field not recognisable.
Colour in preservative: dorsum light brown with poorly defined darker brown patches forming a characteristic pattern: one patch between the eyes, others in dorsolateral position. Weakly developed crossbands on fore- and hindlimbs. A few small white spots scattered on the flanks and on the upper jaw where two rows of 2–3 spots are recognisable, one along the tympanum and directed posteroventrally, the other at mid-eye, directed anteroventrally. Venter beige, throat and chest with brownish pigment forming a rather irregular mottling.
The lower jaw is ventrally marked by an alternating series of contrasting dark and light markings. Colour in life not recorded.
Variation.—Variation in measurements is given in Table 10. See Fig. 70 View FIGURE 70 for colouration in life and its variation. A light vertebral line can be present. There may be weak sexual size dimorphism, but our sample size is small (confirmed male SVL 21.0 mm [n = 1] vs confirmed female SVL 25.1 mm [n = 1]). This is confirmed by additional measurements taken in the field from two males (both 23.5 mm) and three females (27.0– 29.3 mm). Male and female relative tympanum size seems to be similar (HTD/ED ratio is 91% in the female, 88% in the male). Femoral glands of males in life not documented; in one female, rudimentary glands are visible ( Fig. 70d View FIGURE 70 ).
Natural history.—Can be observed during day and night. At Ambatovory, specimens were found in a primary rainforest area characterized by a slow-moving stream and extended forested swamp areas next to the running water; calls were emitted from near-stagnant areas in this swamp. Often sitting in shallow parts of slow-moving streams or on the shore. Occurs in syntopy with M. katae at Talatakely, Ranomafana National Park, and at a stream with some riparian vegetation in a degraded part close to a forest fragment and banana plantation at Andalangina.
Found at an elevational range between 450–971 m a.s.l. in Andalangina and Talatakely. A female with visible eggs was detected on the bank of a slow-moving stream with shallow parts at Talatakely on 6 May 2010, two females with visible eggs were found at Andalangina, one on 25 May 2010, and one on 25 May 2011.
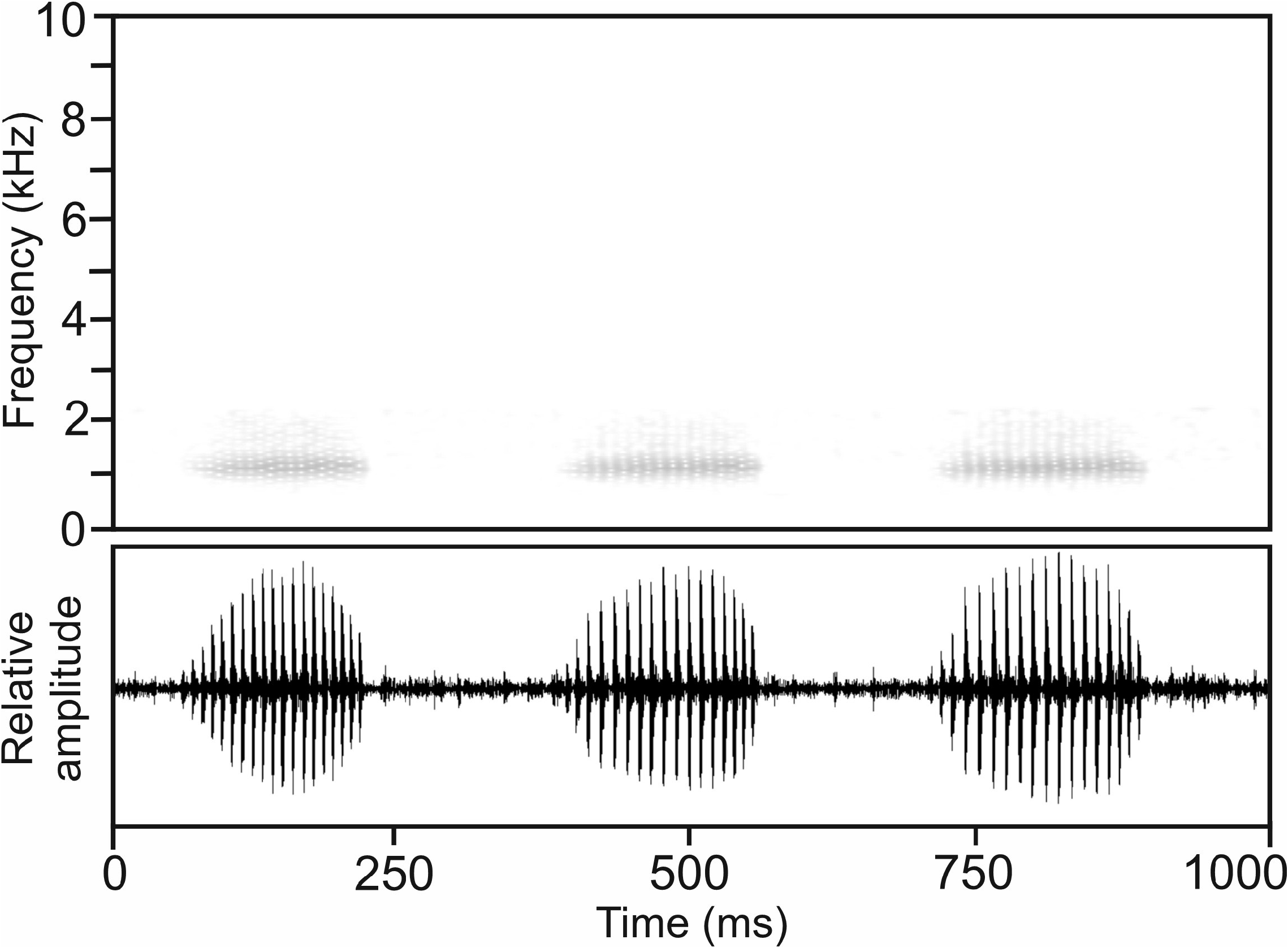
Calls.—A single call series containing six calls recorded on 26 February 2006 at Ambatovory, air temperature not taken, is here assigned to M. glosi , although this assignment is somewhat uncertain given that the calling male has not been observed. The advertisement call consisted of a short, regularly pulsed note repeated at regular intervals and fast succession ( Fig. 71 View FIGURE 71 ). Notes exhibit some amplitude modulation with a fast increase of call energy at the beginning of the note, reaching its maximum approximately at half of the note’s duration.
Call energy was distributed in a relatively narrow frequency band. Numerical parameters of six analysed calls were as follows: call duration (= note duration) 159–214 ms (190.8 ± 20.7 ms); 18–23 pulses per note (20.3 ± 2.3); pulse duration 4–6 ms (4.9 ± 0.5); pulse repetition rate within notes 90.9–115.4 pulses/s (104.1 ± 10.4); dominant frequency 1070–1201 Hz (1124 ± 47 Hz); prevalent bandwidth 800–2100 Hz; call repetition rate (= note repetition rate) within regular series ca 180 calls/min.
Tadpoles.— The tadpole of M. glosi was described under the name ‘ M. sp. aff. biporus “Ranomafana”’ by Schmidt et al. (2009).
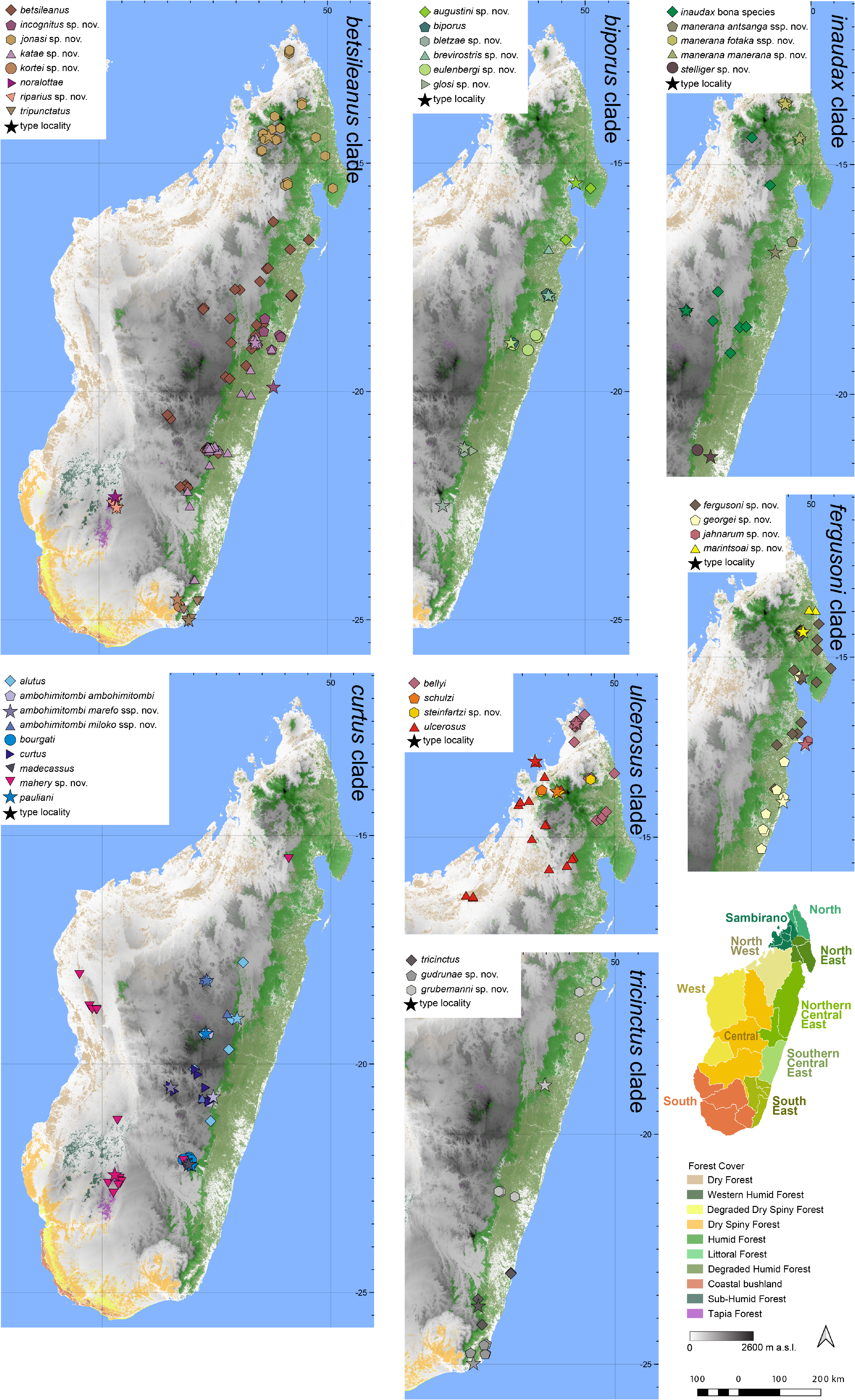
Distribution.— Apparently microendemic to the Ranomafana area ( Fig. 7 View FIGURE 7 ). This species is known from various sites within Ranomafana National Park, and from Andalangina (ca 20 km from Ranomafana). Elevation range: 486–1027 m a.s.l.
Etymology.—We dedicate this species to Julian Glos, in recognition of his contributions to research on conservation of Madagascar’s amphibians.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |