
Mantidactylus stelliger, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504429 |
|
persistent identifier |
https://treatment.plazi.org/id/07B291B4-8CA1-4385-9CCB-106E6AE3DF03 |
|
taxon LSID |
lsid:zoobank.org:act:07B291B4-8CA1-4385-9CCB-106E6AE3DF03 |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus stelliger |
| status |
sp. nov. |
Mantidactylus stelliger sp. nov.
Identity and justification.—This lineage, phenotypically similar to species in the M. biporus clade, occupies an isolated position in the phylogenomic tree and in the 16S tree. It was newly discovered in this study and therefore has not been included in previous DNA barcoding assessments of Madagascar’s amphibians.We here consider it as distinct species due to its isolated phylogenetic position, high mitochondrial divergences of at least 6.8% uncorrected pairwise 16S distance to all other Brygoomantis species, and morphological differences (see Diagnosis below).
Holotype.— ZSM 2381/2007 ( ZCMV 5932 ), adult male, collected by M. Vences, K.C. Wollenberg, and E. Rajeriarison on 3 March 2007 at Ambohitsara (21.3572°S, 047.8157°E), Vatovavy-Fitovinany Region , Madagascar. A 16S barcode sequence of the holotype was obtained in this study and was included in the analysis. GoogleMaps
Paratypes.—A total of four paratypes: ZSM 2379/2007 ( ZCMV 5870 ), adult female, and ZSM 2380/2007 ( ZCMV 5922 ), adult male, with the same collection data as the holotype GoogleMaps ; UADBA uncatalogued ( ZCMV 5923 , ZCMV 5931 ), two specimes of unknown sex and maturity, with the same collection data as the holotype GoogleMaps .
Diagnosis.— Mantidactylus stelliger sp. nov. is the sole member of the M. stelliger clade according to our phylogenomic analysis. See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of small body size (male SVL 21–23 mm, female SVL 31 mm), slightly granular dorsal skin without dorsolateral ridges, large tympanum size in males (12% of SVL), presence of white spots on flanks in at least some individuals, and absence of a white marking on the snout tip, distinguishes M. stelliger sp. nov. from species of the M. betsileanus , M. curtus , M. fergusoni , M. tricinctus , and M. ulcerosus clades. M. inaudax ( M. inaudax clade) differs by larger body size and more developed foot webbing. Species of the M. biporus clade differ as follows: M. biporus by larger body size and and more developed foot webbing; M. augustini by longer hindlimbs and more developed foot webbing; M. bletzae by more developed foot webbing and presence of dorsolateral ridges; M. brevirostris possibly by somewhat smoother dorsal skin and smaller femoral glands; M. eulenbergeri by more developed foot webbing and smoother dorsal skin; M. glosi by shorter hindlimbs and more developed foot webbing ( Table 4 View TABLE 4 ). For a distinction from new species in the M. inaudax clade, see the diagnoses in the respective species accounts below. A full list of molecular diagnostic sites in the 16S gene of M. stelliger sp. nov. in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Description of the holotype. —Adult male in good state of preservation ( Fig. 62 View FIGURE 62 ). Part of right thigh muscle removed as tissue sample. Femoral glands partly detached for examination in internal view. Body rather stout. Head wider than body. Snout rounded in dorsal view. Nostrils directed laterally, not protuberant. Nostrils nearer to tip of the snout than to eye. Canthus rostralis not clearly recognisable. Loreal region very weakly concave.
Tympanum distinct, rounded, its horizontal diameter about 67% of eye diameter. Supratympanic fold present, beginning straight, and gently curving midway towards jaw/forelimb insertion, following the rounded form of the tympanum. Tongue ovoid and bifid. Maxillary teeth present. Vomerine teeth present in two small rounded aggregations, positioned posterolateral to choanae.
Choanae more or less rounded, somewhat elliptical/ slit-like. Subarticular tubercles single. Inner and outer metacarpal tubercles present. Fingers without webbing.
Relative length of fingers: I<II<IV<III. Finger discs minimally enlarged. Nuptial pads absent. Foot of similar length as tibia (99%). Lateral metatarsalia separated. Inner metatarsal tubercle present, outer metatarsal tubercle not clearly recognisable. Webbing formula: 1(1), 2i(1.5), 2e(1), 3i(2), 3e(1.5), 4i(2.5), 4e(3), 5(1.5). Relative length of toes: I<II<V<III<IV. Skin on the upper surface slightly granular in preservative, especially on flanks; in life similar. No dorsolateral folds are recognisable. Ventral side smooth. Femoral glands present, with a distinct and relatively large distal ulcerous macrogland internally consisting of 4–5 large granules. Proximal granular gland field not recognisable.
Colour in preservative: upper surfaces brown with a larger darker brown marking covering much of the anterior dorsum, and areas of lighter grey-brown on the anterior head and posterior dorsum, flanks and limbs.
The whole dorsal surface is covered with numerous poorly defined lighter spots. Dark crossbands on limbs.
On the flanks, several more clearly defined light spots are present, and small white spots also form on the upper jaw a posteroventrally directed row running along the border of the tympanum, and an anteroventrally directed row underneath the eye. Venter light beige, throat and chest brown with light markings and a discontinuous central white line. Colour in life was similar to colour in preservative, with overall more vivid and contrasted pattern. Ventral skin was somewhat translucent, spots on throat and chest silvery white.
Variation.—Variation in measurements is given in Table10. See Fig. 72 View FIGURE 72 for colouration in life and its variation. There may be pronounced sexual size dimorphism, but our sample size is small (confirmed male SVL 21.6–22.6 mm [n = 2] vs confirmed female SVL 31.1 mm [n = 1]). Male and female relative tympanum sizes do not seem to differ (HTD/ED ratio is 74% in the female, 67–74% in the males). Femoral glands of males in life relatively distinct, with a yellowish tone, and mostly consisting of a distinct distal ulcerous macrogland, with only small remnants of a proximal granular gland field.
Natural history.—Poorly known. Specimens were collected from near a stream in a remnant of primary rainforest. At Ranomafana National Park specimens were found during day and night at two streams in primary forest (elevational range between 777–835 m a.s.l.) sitting in shallow parts of the streams. Two females with visible eggs were detected at Sahalavakely on 22 February 2011, another female with visible eggs was found at Sahalavabe on 23 March 2011.
Calls.— The call of this species has not been recorded.
Tadpoles.— The tadpole of this species has not been described.
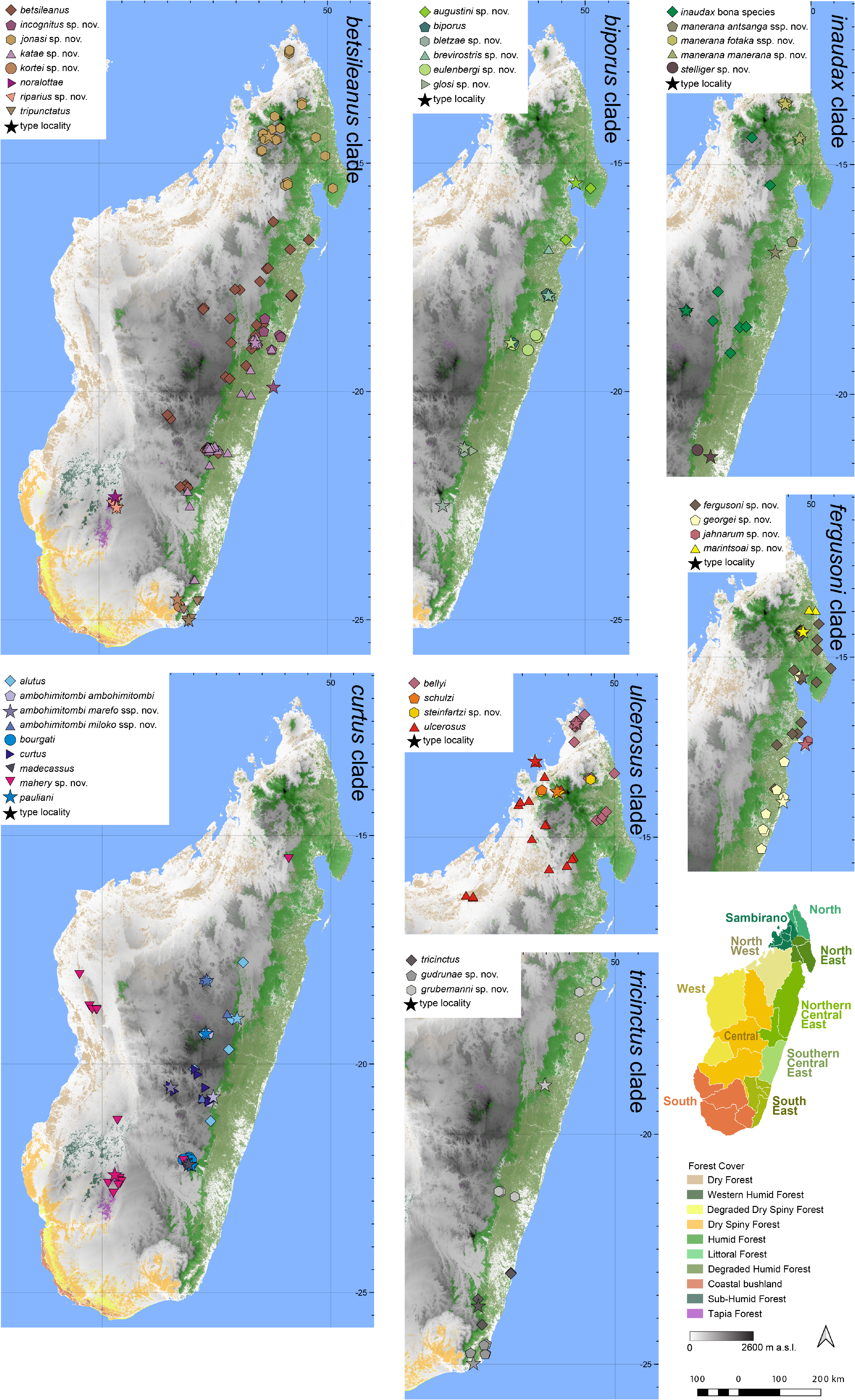
Distribution.— Endemic to the Southern Central East ( Fig. 7 View FIGURE 7 ). This species is known from Ambohitsara and Ranomafana National Park (Sahalavabe and Sahalavakely near Beremby). Elevation range: 294–860 m a.s.l.
Etymology.—The Latin adjective in the masculine nominative singular stelliger, meaning ‘starry’, in reference to the white spots that are often present on the flanks of this species.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |