
Mantidactylus manerana manerana, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504439 |
|
persistent identifier |
https://treatment.plazi.org/id/1C283BB5-F7BA-4D63-807E-9778DF061F12 |
|
taxon LSID |
lsid:zoobank.org:act:1C283BB5-F7BA-4D63-807E-9778DF061F12 |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus manerana manerana |
| status |
subsp. nov. |
Mantidactylus manerana manerana ssp. nov.
Identity and justification.—The nominal subspecies was previously considered as unconfirmed candidate species M. sp. 16 by Vieites et al. (2009), and M. sp. Ca16 by Perl et al. (2014). It was depicted as ‘ Mantidactylus sp. aff. biporus “Andranofotsy”’ by Glaw and Vences (2007).
Holotype.— ZSM 500/2016 (field number ZCMV 15162), adult male, collected by M.D. Scherz, A. Rakotoarison, M. Bletz, M. Vences, and J. Razafindraibe on 17 November 2016 at Camp 3 ‘Simpona’, Marojejy National Park (14.43661°S, 049.74335°E, 1325 m a.s.l.), Sava Region, Madagascar. GoogleMaps A 16S barcode sequence of the holotype was obtained in this study and was included in the analysis.
Paratypes.—A total of 16 paratypes: ZSM 209/2005 ( FGZC 2881 ), adult male, ZSM 206/2005 ( FGZC 2837 ) and ZSM 5064/2005 ( ZCMV 2019 ), two adult females, collected by F. Glaw, M. Vences, and R.D. Randrianiaina on 16–17 February 2005 at the type locality; ZSM 208/2005 ( FGZC 2847 ) and ZSM 207/2005 ( FGZC 2846 ), two adult males, collected by F. Glaw, M. Vences, and R.D. Randrianiaina on 16 February 2005 above Camp 3 ‘Simpona’, Marojejy National Park ; UADBA uncatalogued ( ZCMV 2017–2018 , FGZC 2854 ), three unsexed specimens, UADBA uncatalogued ( FGZC 2835 ), subadult, and UADBA uncatalogued ( FGZC 2836 ), female, collected by F. Glaw, M. Vences, and R.D. Randrianiaina on 16–17 February 2005 around the type locality; UADBA uncatalogued ( ZCMV 2087 ), male, collected by F.Glaw, M.Vences, and R.D. Randrianiaina on 18 February 2005 on the trail between Camp 1 ‘Mantella’ and Camp 2 ‘Marojejia’, Marojejy National Park (precise coordinates unavailable); UADBA uncatalogued ( ZCMV 15190 , ZCMV 15301 , ZCMV 15302 ), three females, UADBA uncatalogued ( ZCMV 15297 ), unsexed adult, UADBA uncatalogued ( ZCMV 15303 ), subadult, with the same collection data as the holotype.
Diagnosis.—See diagnosis for the species, M. manerana , above; for distinction from the other two subspecies, see their diagnoses below.
.
Description of the holotype.—Adult male in excellent state of preservation ( Fig. 73 View FIGURE 73 ). Tissue from left thigh removed. Body rather stout. Head as wide as body. Snout rounded in dorsal view. Nostrils directed laterally, not protuberant, nearer to tip of snout than to eye. Canthus rostralis weakly recognisable, slightly concave; loreal region slightly concave. Tympanum distinct, large, wider than high, horizontal diameter of tympanum 85% of horizontal eye diameter. Supratympanic fold distinct, beginning straight above, with gentle 45° bend midway towards insertion of forelimb. Tongue ovoid, distinctly bifid posteriorly. Maxillary teeth present. Vomerine teeth form two rounded aggregations, positioned posterolateral to choanae. Choanae rounded. Subarticular tubercles single. Inner and outer metacarpal tubercles present. Fingers without webbing. Relative length of fingers: I<II<IV<III. Finger discs slightly enlarged.
Nuptial pads absent. Foot slightly longer than tibia (108%). Lateral metatarsalia separated. Inner metatarsal tubercle present. Outer metatarsal tubercle very small but recognisable. Webbing formula: 1(1), 2i(1.5), 2e(1), 3i(2), 3e(1.5), 4i(2.25), 4e(2.25), 5(0.75). Relative length of toes: I<II<V<III<IV. Skin on the upper surface smooth, slightly glandular dorsolaterally. Two weakly expressed, discontinuous dorsolateral ridges on anterior part of dorsum. Ventral side smooth. Femoral glands small but distinct in external view.
Colour in preservative: dorsally brown, with a dark patchposteriorlybetweeneyeswhichanteriorlyisbordered by a light band. Weakly defined light dorsolateral bands. Numerous distinct contrasted white spots on flanks and laterally on head. An anteroventrally directed white line on the upper lip starts mid-eye. A small white patch on the tip of the snout. Fingers and toes with alternating pattern of light and dark colour. Relatively distinct dark crossbands on limbs. Ventrally beige, with rather contrasted brown pigmentation on throat and chest with light spots and vermiculations, including a median light line on throat, and alternating light-dark pattern ventrally on lower lip. In life, colour was similar to the state in preservative but in general more vivid and contrasted. The white spots on flanks were very contrasted. The lighter parts on the dorsum were light brown. Ventrally silvery white except the darker areas on throat and chest. Femoral glands with a very slight yellow-orange shade.
Variation.—Variation in measurements is given in Table 11. See Fig. 76 View FIGURE 76 for colouration in life and its variation. There is weak sexual size dimorphism (confirmed male SVL 25.0– 27.5 mm [n = 4] vs confirmed female SVL 28.0– 29.4 mm [n = 2]). Males appear to have only slightly larger tympanum sizes than females (HTD/ED ratio is 60–78% in females, 67–86% in males).
Femoral glands of males are relatively small but distinct in life, with a yellowish or orange shade, and with both a distal ulcerous macrogland and a proximal granular gland field recognizable.
Natural history.—Poorly known. Calling specimens were collected at dusk from a stream in primary rainforest.
Calls. —The advertisement call of M. manerana manerana , recorded from the holotype on 17 November 2016, 17:20 h, at Camp Simpona, Marojejy National Park, ca 20°C air temperature, consisted of a pulsed note, emitted isolated or in short series ( Fig. 77 View FIGURE 77 ). Notes exhibit amplitude modulation, with call energy constantly increasing from the beginning of the note, reaching its maximum after approximately one third of the note’s duration. Pulse repetition rate within notes was highest at the beginning and decreased towards the note’s end.
Numerical parameters of eight analysed calls were as follows: call duration (= note duration) 260–363 ms (292.2 ± 37.4 ms); 26–31 pulses per note (27.9 ± 1.8); pulse duration 3–5 ms (4.5 ± 0.7 ms); pulse repetition rate within notes 62.5–115.9 pulses/s (93.9 ± 15.6); dominant frequency 1452–1539 Hz (1498 ± 30 Hz); prevalent bandwidth 720–5300 Hz; call repetition rate (= note repetition rate) within series ca 30 calls/min.
Tadpoles.— The tadpole of M. manerana manerana was described under the name ‘ M. sp. aff. biporus “Marojejy”’ by Schmidt et al. (2009).
Distribution.— Microendemic to the Marojejy massif ( Fig. 7 View FIGURE 7 ). This subspecies is known exclusively from high elevation in Marojejy National Park (from Camp 2 ‘Marojejia’ to above Camp 3 ‘Simpona’). Elevation range: 615–1576 m a.s.l.
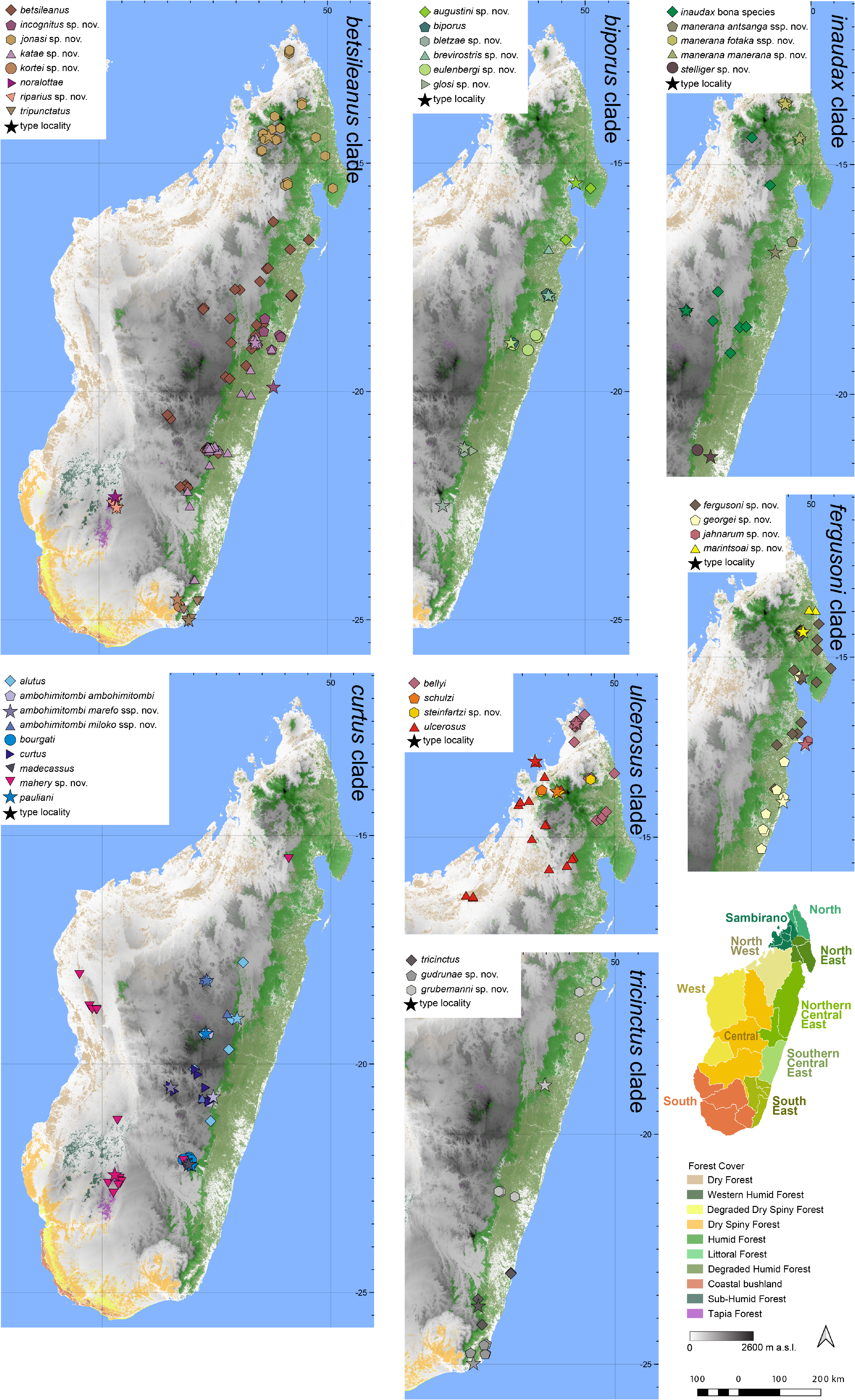


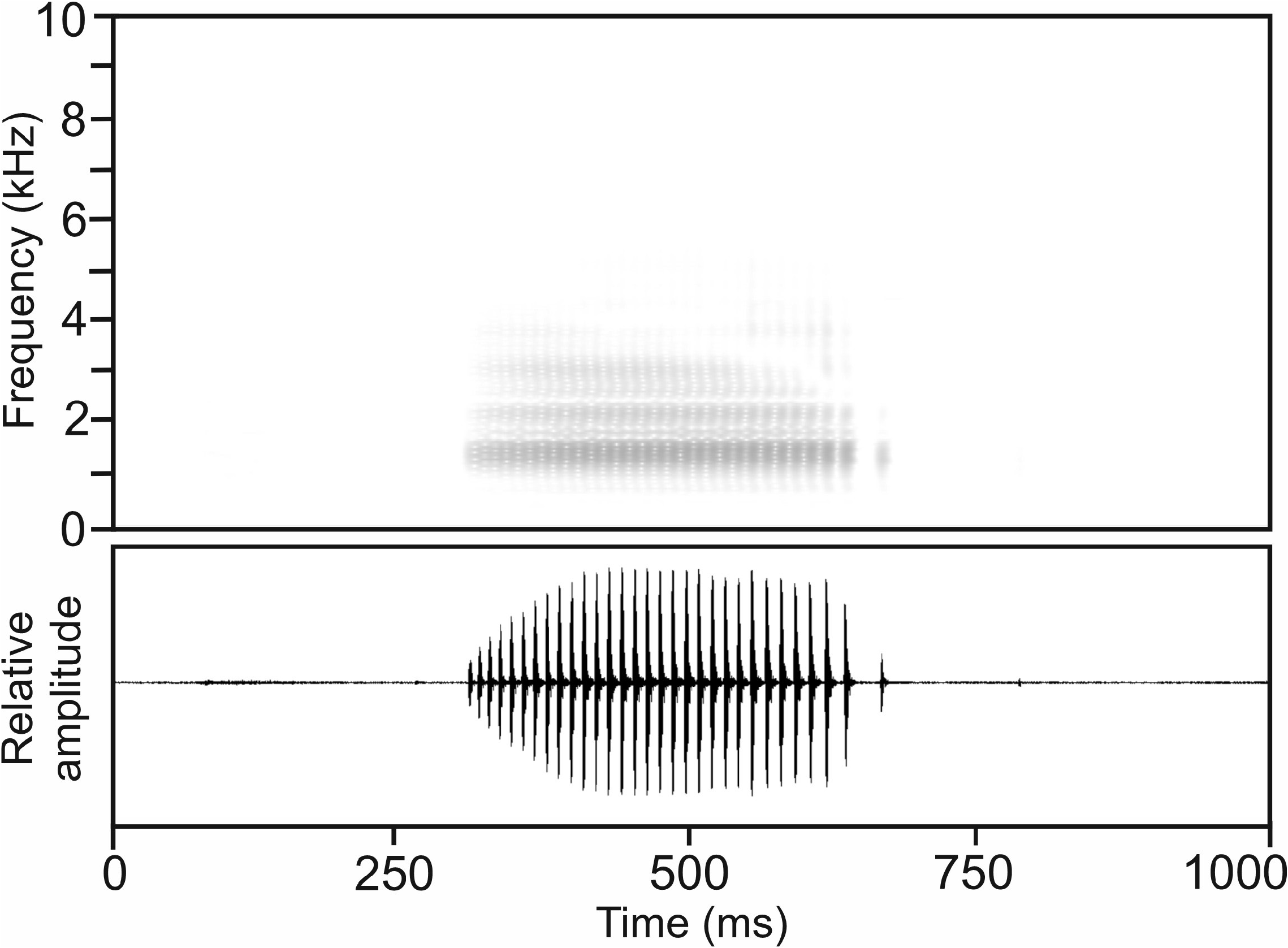
TABLE 11. Morphometric measurements (all in mm) of voucher specimens of the Mantidactylus inaudax clade. Type status is given in square brackets after voucher number: HT,
| Voucher | Field number | Sex | Locality | SVL | HW | HL | ED | HTD | END | NSD | NND | FORL | HAL | HIL | FOTL | FOL | TIBL | FGL | FGW |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mantidactylus inaudax bona species (Ca17) | |||||||||||||||||||
| MZUT An727.1 [LT*] | NA | F | 32.1 | 12.7 | 12.4 | 4.9 | 3.5 | 2.2 | 2.0 | 3.3 | 17.7 | 8.5 | 47.9 | 21.8 | 15.4 | 13.5 | NA | NA | |
| ZSM 180/2005 | FGZC 2146 | M | Ambohitantely | 26.2 | 10.2 | 10.5 | 3.7 | 4.1 | 1.9 | 1.8 | 2.6 | 14.1 | 6.8 | 34.4 | 16.6 | 11.8 | 10.2 | 4.5 | 1.7 |
| ZFMK 60142 # | NA | F | Ambohitantely | 26.9 | 11.8 | 9.4 | 3.8 | 2.6 | 2.2 | 2.1 | 3.2 | 9.1 | 7.8 | NM | NM | 11.0 | 11.0 | NA | NA |
| ZMA 19310 # | FGMV 2002.2435 | F | Fierenana | 33.1 | 13.3 | 12.7 | 5.2 | 3.3 | 2.7 | 2.1 | 4.1 | 11.3 | 9.1 | NM | NM | 14.6 | 14.6 | 1.6 | 1.3 |
| ZMA 19311 # | FGMV 2002.2439 | F | Fierenana (pitfall forest) | 27.4 | 11.3 | 9.9 | 3.8 | 3.1 | 2.3 | 1.9 | 3.3 | 11.4 | 8.6 | NM | NM | 13.8 | 13.6 | 1.6 | 1.4 |
| ZMA 19314 # | FGMV 2002.2429 | F | Fierenana (Camp) | 27.5 | 11.5 | 9.9 | 4.0 | 3.0 | 2.1 | 1.7 | 3.0 | 10.9 | 8.6 | NM | NM | 14.0 | 14.5 | 1.2 | 1.2 |
| ZMA 19331 # | FGMV 2002.2441 | F | Fierenana (pitfall forest) | 27.0 | 11.1 | 9.6 | 3.8 | 3.0 | 2.1 | 1.9 | 3.2 | 11.4 | 8.8 | NM | NM | 13.6 | 13.7 | 1.5 | 1.2 |
| ZSM 181/2005 | FGZC 2147 | F | Ambohitantely | 29.4 | 11.0 | 11.2 | 4.0 | 3.1 | 2.3 | 1.8 | 2.8 | 15.5 | 7.5 | 40.2 | 19.3 | 13.1 | 11.6 | NA | NA |
| ZMA 19341 # | FGMV 2002.2252 | F? | Fierenana (Camp) | 33.6 | 12.9 | 11.5 | 4.7 | 3.6 | 2.4 | 1.9 | 3.8 | 11.9 | 9.9 | NM | NM | 15.7 | 15.4 | 1.7 | 1.2 |
| ZFMK 60141 # | NA | M | Ambohitantely | 21.7 | 9.8 | 7.8 | 3.2 | 2.9 | 1.7 | 1.7 | 2.5 | 8.5 | 6.4 | NM | NM | 9.7 | 10.1 | 2.2 | 1.6 |
| ZMA 19343 # | FGMV 2002.2423 | M | Fierenana (Camp) | 27.4 | 11.0 | 9.6 | 3.7 | 3.4 | 2.1 | 1.8 | 3.3 | 10.0 | 8.7 | NM | NM | 12.5 | 12.3 | 2.7 | 2.0 |
| ZMA 19345 # | FGMV 2002.2251 | M | Fierenana (Camp) | 25.8 | 10.5 | 9.4 | 3.8 | 3.2 | 2.1 | 1.7 | 3.0 | 9.5 | 7.6 | NM | NM | 11.3 | 11.0 | 3.0 | 2.2 |
| ZSM 1768/2008 | ZCMV 8869 | M | Forest fragment Brieville | 25.7 | 10.6 | 10.5 | 4.0 | 3.6 | 2.3 | 1.8 | 3.2 | 15.0 | 6.7 | 38.2 | 17.6 | 12.1 | 10.4 | 2.7 | 1.9 |
| ZSM 363/2010 | FGZC 4358 | M | Fanamby Forest, Anjozorobe | 23.7 | 9.0 | 9.1 | 3.4 | 3.4 | 2.2 | 1.6 | 2.3 | 13.0 | 6.8 | 31.6 | 15.2 | 10.7 | 9.4 | 2.9 | 2.1 |
| ZSM 398/2006 | ZCMV 3259 | M | Moramanga- Anosibeanala | 30.0 | 10.7 | 11.9 | 4.3 | 3.6 | 2.3 | 2.1 | 3.3 | 15.8 | 8.3 | 42.2 | 20.7 | 14.5 | 11.5 | 2.8 | 2.2 |
| ZSM 548/2009 | ZCMV 11213 | M | Makira West | 22.7 | 8.7 | 9.5 | 3.8 | 3.3 | 2.2 | 1.5 | 2.4 | 13.5 | 7.0 | 33.9 | 16.5 | 11.7 | 9.8 | 4.4 | 2.0 |
...Continued on the next page
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |