
Mantidactylus grubenmanni, Scherz & Crottini & Hutter & Hildenbrand & Andreone & Fulgence & Köhler & Ndriantsoa & Ohler & Preick & Rakotoarison & Rancilhac & Raselimanana & Riemann & Rödel & Rosa & Streicher & Vieites & Köhler & Hofreiter & Glaw & Vences, 2022
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504405 |
|
persistent identifier |
https://treatment.plazi.org/id/630AA2B0-064E-4623-BE3B-A28C8657C03C |
|
taxon LSID |
lsid:zoobank.org:act:630AA2B0-064E-4623-BE3B-A28C8657C03C |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus grubenmanni |
| status |
sp. nov. |
Mantidactylus grubenmanni sp. nov.
Identity and justification. —This lineage is one of three species in the M. tricinctus clade. It corresponds to specimens from An’Ala used by Glaw and Vences (1999) for a redescription of M. tricinctus ; however, it is very stronglydivergentinmitochondrialDNAandconcordantly, also in Rag-1 sequences, and differs in advertisement calls, suggesting it represents a distinct species. It was depicted as ‘ Mantidactylus sp. aff. tricinctus “An’Ala”’ by Glaw and Vences (2007).
Holotype.— ZSM 375/2006 ( ZCMV 1404 ), adult male with distinct femoral glands, collected by D.R. Vieites, M. Vences, F. Rabemananjara, P. Bora, C. Weldon, and J. Patton on 7–8 February 2006 at An’Ala (forest camp; 18.91926°S, 048.48796°E, 889 m a.s.l.), AlaotraMangoro Region , Madagascar. A 16S barcode sequence of the holotype is available from GenBank (accession FJ559281 View Materials ). GoogleMaps
Paratypes.—A total of eight paratypes: ZSM 376/2006 ( ZCMV 1425 ), unsexed adult without externally visible femoral glands (probably a female), with the same collecting data as the holotype. GoogleMaps Furthermore, we also designate as paratypes ZFMK 62252–62254 , three adult females, and ZFMK 62251 , 62255–62257 , four adult males, collected by F. Glaw and D.M. Rakotondramanana on 3 February 1996 in An’Ala ; GoogleMaps although the identity of these topotypical specimens was not confirmed by genetic data, their morphological identity is unambiguous.
Additional material.— The following specimens belong to genetically divergent populations and therefore are not included in the paratype series: ZMB 81959 ( JCR 616 ), adult female, collected on 1 March 2011 by J.C. Riemann, and S.H. Ndriantsoa at Ambolo , Ranomafana area (21.26307°S, 047.50696°E, 660 m a.s.l.) GoogleMaps ; ZMB 81960 ( JCR 694 ), adult male, collected on 14 March 2011 by J.C. Riemann, and S.H. Ndriantsoa at Imaloka , Ranomafana area (21.24274°S, 047.46507°E, 1052 m a.s.l.) GoogleMaps ; ZMB 81958 ( JCR 419 ), adult female, collected on 2 June 2010 by J.C. Riemann, and S.H. Ndriantsoa at Antaramanavana , Ranomafana area (21.23997°S, 047.50647°E, 641 m a.s.l.) GoogleMaps ; ZMB 81961 (field NSH 1900, GenBank JCR 1900), collected on 26 March 2011 by J.C. Riemann, and S.H. Ndriantsoa at Antaramanavana , Ranomafana area (21.23997°S, 047.50647°E, 641 m a.s.l.) GoogleMaps ; ZMB 81962 (JCR 874), adult male, collected on 23 May 2011 by J.C. Riemann, and S.H. Ndriantsoa at Ambolo , Ranomafana area (21.26307°S, 047.50696°E, 660 m a.s.l.) GoogleMaps ; ZMB 81963–81964 (JCR 922–923), adult male and female, collected on 9 June 2011 by J.C. Riemann, and S.H. Ndriantsoa at Antaramanavana , Ranomafana area (21.23997°S, 047.50647°E, 641 m a.s.l.) GoogleMaps ; ZSM 2416/2007 ( ZCMV 5925 ) and ZSM 2417/2007 ( ZCMV 5930 ), female and male from Ambohitsara (21.3571°S, 047.8156°E), collected on 3 March 2007 by M. Vences, K.C. Wollenberg, and E. Rajeriarison. GoogleMaps
Diagnosis.— Mantidactylus grubenmanni sp. nov. is a member of the M. tricinctus clade as revealed by the phylogenomic analysis, and sister to M. tricinctus . See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of very small body size (below 20 mm male SVL), connected lateral metatarsalia, reduced webbing, presence of a light (often yellowish) marking on snout tip, a yellow inguinal marking, and a short, pulsed advertisement call emitted in rapid succession in regular series, readily distinguishes M. grubenmanni sp. nov. from all other nominal species of Brygoomantis except its sister species M. tricinctus from which it is morphologically indistinguishable. It differs from M. tricinctus by fewer pulses per note in advertisement calls, and a generally lower pulse repetition rate. For detailed distinction from other new species described herein, see the respective species accounts. A full list of molecular diagnostic sites in the 16S gene of M. grubenmanni sp. nov. in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Description of the holotype. —Adult male in mediocre state of preservation ( Fig. 56 View FIGURE 56 ). Tissue removed from left thigh. Belly cut open for parasitological examination. Left femoral glands partly detached for examination in internal view. Body slender. Head slightly wider than body. Snout rounded in dorsal view, slightly truncate in lateral view. Nostrils directed laterally, slightly protuberant, nearer to tip of snout than to eye. Canthus rostralis weakly recognisable, slightly concave; loreal region concave. Tympanum distinct and large, rounded, horizontal diameter of tympanum 93% of horizontal eye diameter. Supratympanic fold in its first part almost identical to tympanum edge, thereafter distinct, running rather straight from behind eye and bending about 45° close to posterior edge of tympanum towards forelimb insertion. Tongue ovoid, distinctly bifid. Maxillary teeth present. Vomerine teeth form two small, rounded patches, positioned posteromedial from choanae. Choanae rounded. Subarticular tubercles single. Inner and outer metacarpal tubercles present. Fingers without webbing. Relative length of fingers: I<II<IV<III. Finger discs slightly enlarged. Nuptial pads absent. Foot almost of exact same length as tibia (101%). Lateral metatarsalia connected. Inner and small outer metatarsal tubercle present. Feet with very poorly expressed webbing, absent or only traces among some toes. Webbing formula: 1(no web), 2i(no web), 2e(1.5), 3i(2.5), 3e(2), 4i(3), 4e(traces), 5(traces). Relative length of toes: I<II<V<III<IV. Skin on the upper surface smooth with some scattered granules. Ventral side smooth. Femoral glands with a large and distinct distal ulcerous macrogland; no clearly recognisable proximal granular gland field.
Colour in preservative: dorsally almost uniformly brown, with a lighter anterior surface of head and some indistinct lighter markings laterally on head. Limbs with dark crossbands. Ventrally light beige with a small amount of brown marking on chest, anterior belly and posterior part of throat. No clear median light line recognisable on throat. Lower lip ventrally with alternating white / dark brown pattern. Colour of holotype in life not documented.
Variation.—Variation in measurements is given in Table 9. See Fig. 59 View FIGURE 59 for colouration in life and its variation. There is moderate sexual size dimorphism (confirmed male SVL 16.7–17.9 mm [n= 5] vs confirmed female SVL 19.2–20.3 mm [n = 3]). Males have an only slightly larger tympanum than females (HTD/ED ratio is 68–87% in females, 81–93% in males). Femoral glands of males in life quite distinct and coloured with a conspicuous yellowish shade; a large and distinct distal ulcerous macrogland is clearly visible.
Natural history.—As reported by Glaw and Vences (1999), males in An’Ala have been observed calling at daytime in a shallow, partly sun-exposed swamp with dense vegetation in primary forest, near or on perches directly above the water. They were sitting on leaves, fallen branches and similar structures, generally only 0–2 cm above the water level. Calling behaviour of one individual male was observed for several minutes. This specimen moved forward during its vocalisations, in a conspicuously jerky, disrupted way. In the Ranomafana area, found along streams in rainforest inside Ranomafana National Park and forest fragments in Ambolo and Ampitavanana, but not found outside forest. Specimens were observed sitting in areas of shallow water and along the banks or hiding in leaf litter in the vicinity of a stream. At Ranomafana and surrounds found at an elevational range between 641–1052 m a.s.l. A female with visible eggs was found in June 2010 at Ampitavanana, another in March 2011 at Ambolo. ZMB 81960 (JCR 694) was observed calling at a stream bank at Imaloka inside Ranomafana National Park at late afternoon (17:00 h) on 14 March 2011.
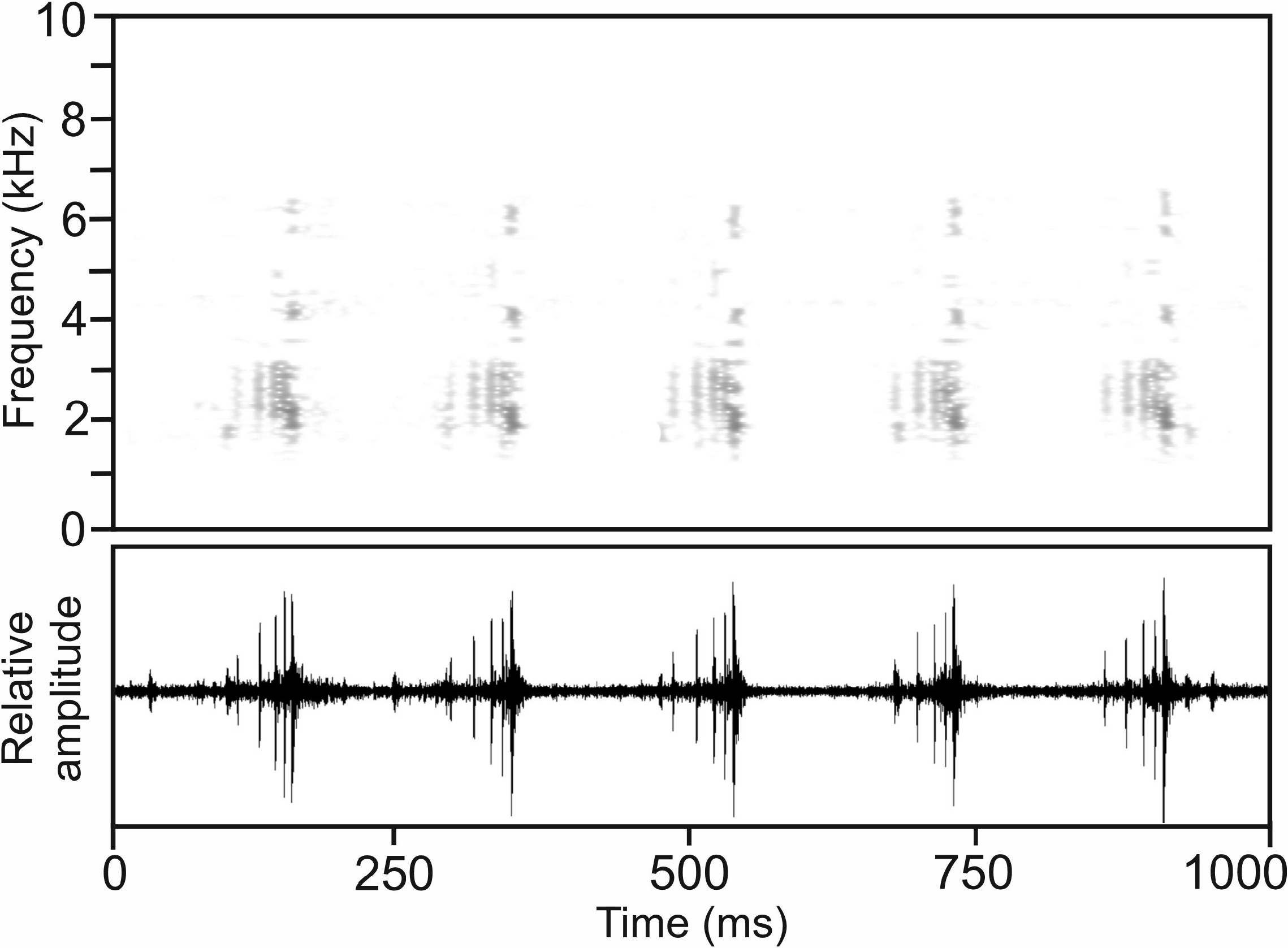
Calls.—The advertisement call of M. grubenmanni , recorded on 3 February 1996 at An’Ala, 29.6°C air temperature ( Vences et al. 2006: CD2, track 71), consisted of a very short, pulsed note, emitted singly or in long regular series at very fast succession ( Fig. 60 View FIGURE60 ). Amplitude modulation was present, with relative amplitude increasing from the beginning of the call, reaching its maximum with the terminal pulse. Pulse structure within notes was somewhat complex, with terminal pulses having longer duration and being more narrowly spaced. Numerical parameters of 52 analysed calls were as follows: call duration (= note duration) 26–87 ms (68.1 ± 17.5 ms); 3–7 pulses per note (5.5 ± 0.9); pulse duration 2–12 ms (4.5 ± 2.8); pulse repetition rate within notes 47.6–181.8 pulses/s (89.2 ± 40.3); dominant frequency 2015–2338 Hz (2286 ± 104 Hz); prevalent bandwidth 1500–3400 Hz; call repetition rate (= note repetition rate) within regular series ca 310 calls/min. The longest call series recorded had a duration of 8.52 s and contained 43 calls. Calls recorded on 14 March 2011 at Ranomafana from specimen ZMB 81960 (JCR 694) agree in structure and parameters with those described above from An’Ala. The longest regular call series from this recording had a duration of 7.15 seconds and contained 33 calls, repeated at a rate of 272 calls/minute.
Tadpoles.— The tadpole of this species has not been described.
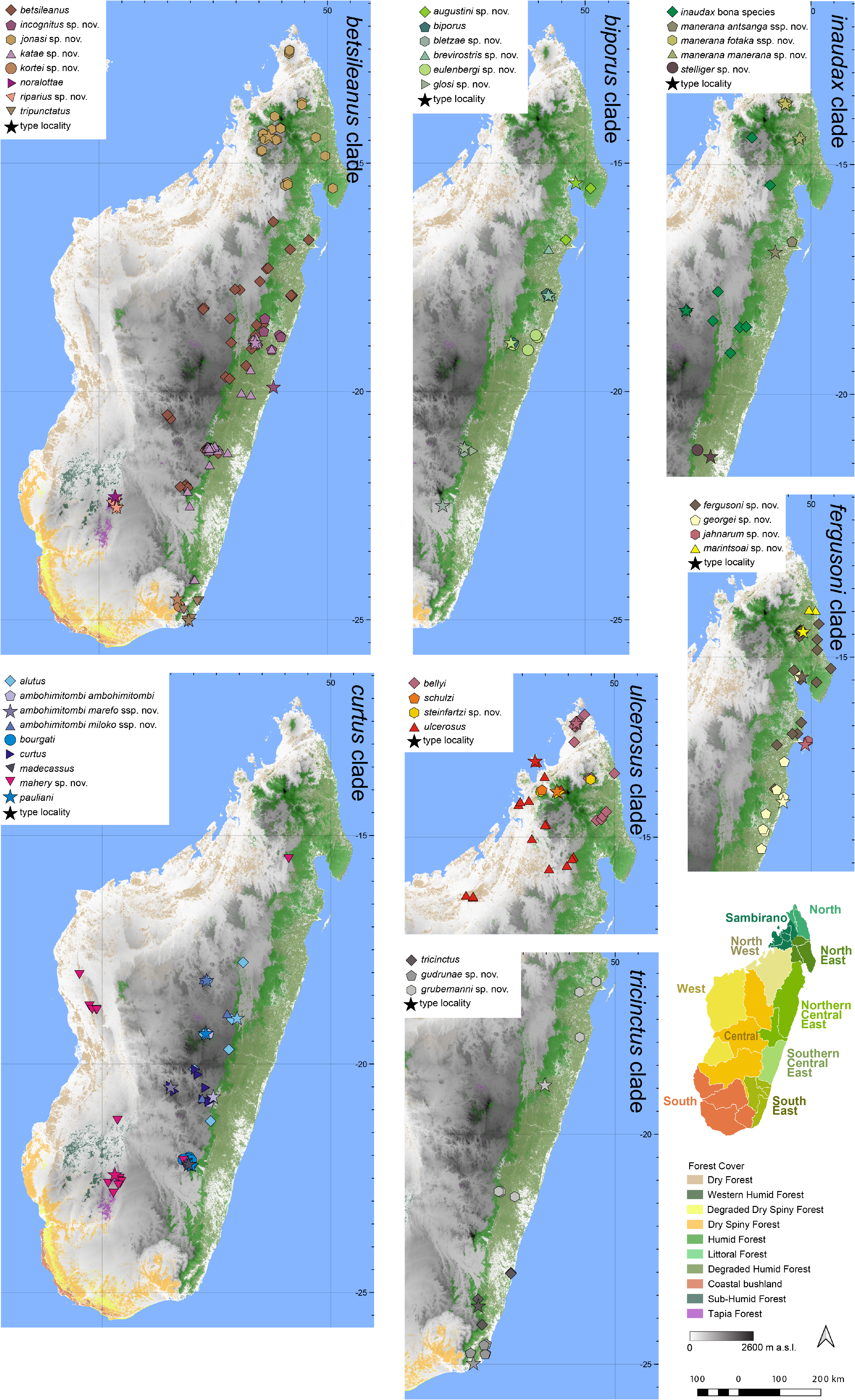
Distribution.— Widespread in Northern Central East and Southern Central East ( Fig. 7 View FIGURE 7 ). This species is known from Ambatobe, Ambohitsara, An’Ala, Befanjana, Betampona, several localities in Ranomafana (Ambolo, Ampitavanana, Beremby, Imaloka), and Sahavontsira. Elevation range: 14–1052 m a.s.l.
Etymology.— We dedicate this species to Moritz Grubenmann, dedicated microbiologist and naturalist from Z̧rich with an enormous knowledge on the Malagasy flora and fauna, in acknowledgement of his constant support of our research with numerous important observations and excellent photographs of Malagasy amphibians and reptiles.


| ZFMK |
Germany, Bonn, Zoologische Forschungsinstitut und Museum "Alexander Koenig" |
| ZMB |
Germany, Berlin, Museum fuer Naturkunde der Humboldt-Universitaet |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |