
Ambopteryx longibrachium, Wang & O’Connor & Xu & Zhou, 2019
|
publication ID |
https://doi.org/ 10.1038/s41586-019-1137-z |
|
DOI |
https://doi.org/10.5281/zenodo.6485756 |
|
persistent identifier |
https://treatment.plazi.org/id/5E0C87B1-7643-696A-FF5C-FA60FE65DCC4 |
|
treatment provided by |
Plazi |
|
scientific name |
Ambopteryx longibrachium |
| status |
gen. et sp. nov. |
Ambopteryx longibrachium gen. et sp. nov.
Etymology. ‘ Ambo ’, both (Latin); ‘ pteryx ’, wing (Latin), referring to the pterosaur-like wing that is present in this non-avian dinosaur; ‘ longibrachium ’ refers to the elongate forelimb (brachium, upper arm (Latin)).
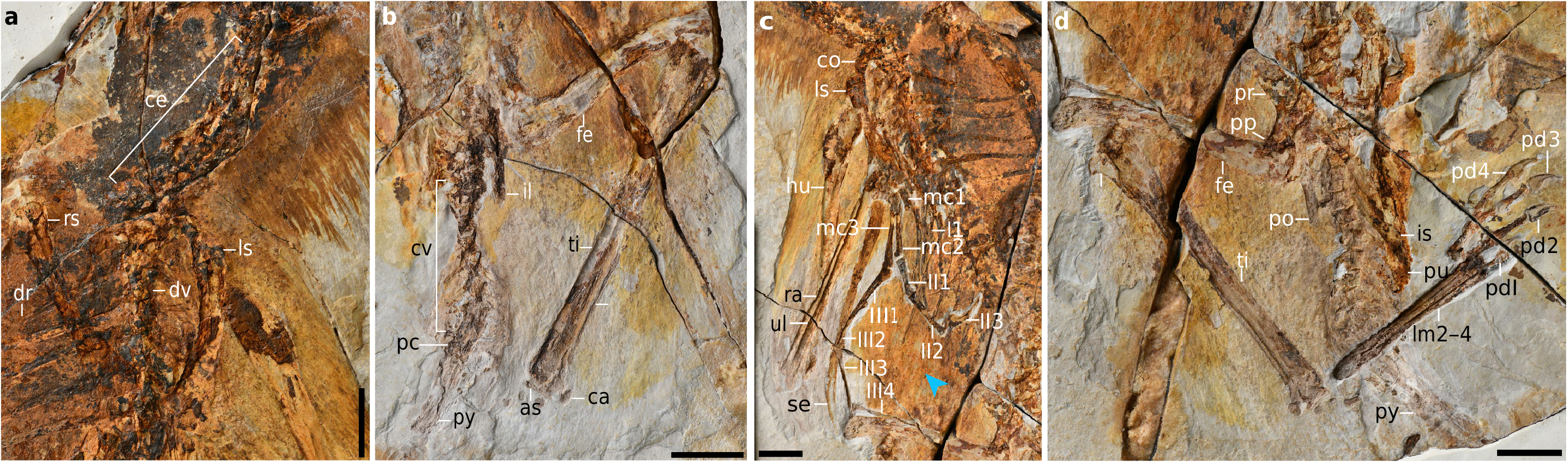
Holotype. Institute of Vertebrate Paleontology and Paleoanthropology ( IVPP) V24192 View Materials , an articulated and nearly complete skeleton with associated soft tissue preserved on a slab and counter slab ( Fig. 1 View Fig , Extended Data Fig. 1 View Fig ).
Locality and horizon. Near Wubaiding Village, Reshuitang Town, Lingyuan City, Liaoning Province, China; early Upper Jurassic, stratigraphic equivalent of the Haifanggou Formation , Yanliao Biota ( Oxfordian stage, 163 million years ago (Ma) 8, 9; see Methods for the provenance of the holotype specimen).
Diagnosis. A scansoriopterygid theropod that preserves the following synapomorphies of the Scansoriopterygidae : scapula distally expanded; reduced deltopectoral crest of humerus; and metacarpal III longer than II, and elongate manual digit III that is longer than other digits. The presence of a rod-like styliform element and short tibia ally the Ambopteryx specimen with the genus Yi, but Ambopteryx is distinguishable on basis of the following features: (1) ulna substantially wider (2×) than radius; (2) short tail that lacks a transition point and that ends in a fused pygostyle; (3) elongate forelimb 1.3× longer than the hindlimb; (4) straight, rod-like postacetabular process of ilium; and (5) tibia shorter than humerus (see Methods for differential diagnosis; the three manual digits of Pennaraptora are here denoted as I, II and III, although the homologies of the manus in tetanuran theropods are debated 3). Features (1), (2) and (4) are probable autapomorphies that are thus far unique to Ambopteryx, but that cannot be compared across all scansoriopterygids.
IVPP V24192 View Materials is interpreted as a subadult or adult on the basis of the fusion of the pygostyle, and the absence of striations or pits to indicate periosteal ossification was incomplete. Osteohistological examination was attempted but no information could be extracted, owing to poor preservation and diagenetic alteration (Extended Data Fig. 2a, b View Fig ). IVPP V24192 View Materials is estimated to be 32 cm in body length and to have had a body mass approximately 306 g, using an empirical equation based on femur circumference 10 (see Methods).
The skull is anteroposteriorly short and dorsoventrally high (Extended Data Fig. 1c View Fig ), similar to other scansoriopterygids 6. The neck is complete and consists of eight-to-ten articulated cervicals (axis and atlas included) ( Fig. 2a View Fig ). As in some oviraptorosaurs and therizinosaurs 11 —but unlike most other non-avian pennaraptorans 12 — the tail is short and ends in a pygostyle.In contrast to most other non-avian theropods 13 and stemward avians 12, the free caudals lack a transition point.
The forelimb is gracile and markedly elongate ( Fig. 2c View Fig , Extended Data Fig. 3c View Fig ), with an intermembral index ((humerus + ulna)/ (femur + tibia)) of about 1.3, which surpasses that of other scansoriopterygids, as well as most non-avian theropods and stem avians (Supplementary Tables 1–3). The proximal margin of the humerus is strongly convex, whereas in Yi it is relatively flat. The straight ulna is shorter than the humerus and is nearly twice the transverse width of the radius, which is in stark contrast to the subtle difference between the ulna and the radius that is present in other scansoriopterygids 6, 14, 15 and many non-avian theropods 16. An elongate bone is preserved that extends from the distal end of the left ulna; this bone is morphologically identical to the anomalous structure reported in Yi 6, being rod-like, slightly curved and tapered distally (Extended Data Fig. 4 View Extended ). No comparable structure has hitherto been found in other dinosaurs 6. We consider this bone homologous to that of Yi, and hereafter use the same terminology in referring to this bone as the styliform element 6. The styliform element is shorter than the humerus in Ambopteryx, which is not the case in Yi. The hand morphology is characteristic of scansoriopterygids in that it shows a metacarpal III that is longer than metacarpal II, and an elongate digit III that is substantially longer than digit II 6, 14 ( Fig. 2c View Fig ). The manual phalangeal formula is 2-3-4. Phalanx I-1 terminates level with the distal end of metacarpal II, whereas in Yi and Epidendrosaurus this phalanx extends further distally (Extended Data Fig. 5 View Extended ).
The postacetabular process of the ilium is dorsoventrally narrow (less than one-third of the height of the preacetabular process) and strap-like in lateral view (Extended Data Fig. 3b, d View Fig ), which contrasts with both the triangular form as in some oviraptorosaurs and troodontids 17, 18 and the ventrally curved condition as in dromaeosaurids and stem avians 19. As in Yi, the tibia is shorter than the humerus; the opposite is true in other non-avian pennaraptorans (Supplementary Table 3). Ambopteryx preserves approximately 20 stones and unidentified bony fragments just anterior to the pelvis and ventral to the vertebral column, which are interpreted as gastroliths and ingested remains, respectively ( Fig. 1 View Fig , Extended Data Fig. 1d View Fig ). The diet of scansoriopterygids has been unknown; the gastroliths and possible stomach contents preserved in Ambopteryx provide the some direct clues from which we can draw dietary inferences. The small number of gastroliths in Ambopteryx suggests that this taxon may not have been herbivorous—this also finds some support in the stomach contents, which appear to contain bone. Considering the unusual dental morphology of the scansoriopterygid clade, we suggest this group may have been omnivorous.
Feathers are densely preserved around the head, neck and the shoulder regions ( Fig. 1 View Fig , Extended Data Fig. 1 View Fig ). The matrix that surrounds the left hand, right forelimb and abdomen is coated in a brownish continuous layer that bears ripple-like striations ( Fig. 2c View Fig ), which is reminiscent of the membranous tissues reported in Yi 6. Scanning electron microscopy reveals densely packed microscopic elliptic moulds within the neck feathers and membranous tissues (Extended Data Fig. 6 View Extended ). Following recent studies 20, these moulds are interpreted as being produced by eumelanosomes (see Supplementary Information for additional description and discussion of diet).
Our phylogenetic analysis recovered Ambopteryx as the sister taxon to Yi within the clade Scansoriopterygidae ( Fig. 3a View Fig , Extended Data Fig. 7 View Fig ). The nearly complete postcranial skeleton of Ambopteryx provides a great deal of important morphological and ecological information that is not accessible from previously known scansoriopterygid specimens. The fused pygostyle in Ambopteryx adds to the known distribution of this feature among non-avian dinosaurs. The abbreviated tail and elongate forelimbs in scansoriopterygids—which evolved in parallel similar morphologies in Aves—would have increased stability during possible volant behaviour by shifting the centre of weight more anteriorly, closer to the centre of lift 7.
The presence of a styliform element in Ambopteryx lends further credibility to previous interpretations of this structure in Yi 6 and confirms that this element is a genuine feature. Comparable elements (whether homologous or not) are otherwise unknown among dinosaurs 6, but have evolved independently in some other tetrapod lineages that are adapted for volant behaviour 21, 22. Where the styliform element is present in extant vertebrates, it supports an expanded patagium and thus increases the viable area of the aerodynamic surface 21, 22. Yi and Ambopteryx preserve membranous tissue in situ around the styliform element 6, which indicates a similar patagium-supporting role for this structure in this clade. Unfortunately, the preservation in Ambopteryx and Yi makes it impossible to reconstruct the configuration of the membranous tissue, which prevents further assessment of its exact function. Although previously a gliding function was ascribed to these structures in Yi 6, other functions are also possible—for example, in display and ornamentation, or in contributing to wing-assisted incline running 23. The hyper-elongate manual digit III of scansoriopterygids was previously interpreted as a feeding adaptation for extracting grubs during foraging, using a strategy similar to that of the extant aye-aye 15. This interpretation is not supported in light of new data that indicate that the manual digits were surrounded by membranous tissues 6, and in light of the limited mobility that we inferred from the morphology of the manual phalangeal joints. We suggest the primary function of the elongated digit III was the attachment of the wing membrane rather than feeding behaviour.
Scansoriopterygids truly are one of the most bizarre clades of nonavian theropods. In addition to their membranous wings, their body plan is unique among theropods. Morphometric analyses indicate that scansoriopterygids occupy a distinct morphospace that is separated from most other non-avian coelurosaurians and Mesozoic avians, which suggests that substantial changes in limb proportions evolved after scansoriopterygids diverged from the rest of Paraves ( Fig. 3b, c View Fig , Methods). The forelimb underwent considerable elongation in scansoriopterygids (paralleling changes in other volant paravians), and achieved proportions that are only otherwise observed in Aves. Scansoriopterygids have unusually short metacarpals—the elongation of their forelimb is primarily achieved through the elongation of the humerus and ulna. By contrast, the forelimb elongation in nonscansoriopterygid pennaraptorans (including birds) is largely driven by the elongation of the metacarpals. The presence of an elongate manual digit III and a styliform element in scansoriopterygids probably compensate for the relatively short metacarpals by providing support for membranous wings. By contrast, selection for relatively elongate metacarpals in most paravians may have been driven by the need for increased area for the attachment of the primary remiges (for example, over half of the primary remiges attach to the carpometacarpus in living birds 24). The co-occurrence of short metacarpals with membranous wings—as opposed to long metacarpals and feathered wingsdemonstrates how these two markedly different flight apparatuses can affect the overall forelimb structure. Membranous wings are unknown among Cretaceous theropods, which indicates that the combination of an avian-like body plan (for example, elongate forelimbs and a short tail) with membranous wings that evolved in the Scansoriopterygidae represents a previously unknown adaptation of vertebrates with volant behaviour. Feathered wings, which are first documented in some Upper Jurassic non-avian theropods, were further refined through the evolution of numerous other musculoskeletal modifications and eventually gave rise to the current success of modern birds, which represent the most-diverse extant clade of tetrapods.





| IVPP |
IVPP |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.