
Tellervotrema beringi ( Mamaev, 1965 ) Gibson & Bray, 1982
|
publication ID |
https://doi.org/10.11646/zootaxa.3986.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:B84A49B3-F5F3-44AF-B270-038D6D28A4A2 |
|
DOI |
https://doi.org/10.5281/zenodo.5670170 |
|
persistent identifier |
https://treatment.plazi.org/id/5D5A5F74-0E73-FFC1-B3AD-823E6FA768C9 |
|
treatment provided by |
Plazi |
|
scientific name |
Tellervotrema beringi ( Mamaev, 1965 ) Gibson & Bray, 1982 |
| status |
|
Tellervotrema beringi ( Mamaev, 1965) Gibson & Bray, 1982 View in CoL
( Figs. 4–6 View FIGURES 4 – 6 )
Synonyms. Plagioporus beringi Mamaev, 1965 ; Tellervotrema katadara of Kuramochi (2009).
Host. Coryphaenoides longifilis Günther ; Gadiformes : Macrouridae : Macrourinae ; longfin grenadier.
Locality. Off the Pacific coast of the Tōhoku region, northern Honshu, Japan, 37°47.60’N, 142°37.12’E – 37°47.4’N, 142°37.17’E (Station WA-05-F1200), depth = 1,196 m, 28/October/2005.
Site. Intestine.
Deposited Specimens. Collector TK, 8 vouchers NSMT-Pl 5672.
Records. 1. Mamaev (1965); 2. Yamaguti (1971); 3. Gibson & Bray (1982); 4. Bray (1995); 5. Blend (1996); 6. Blend et al. (2012); 7. Kuramochi (2009); 8. Present study.
Descriptions. 1, 6, 8.
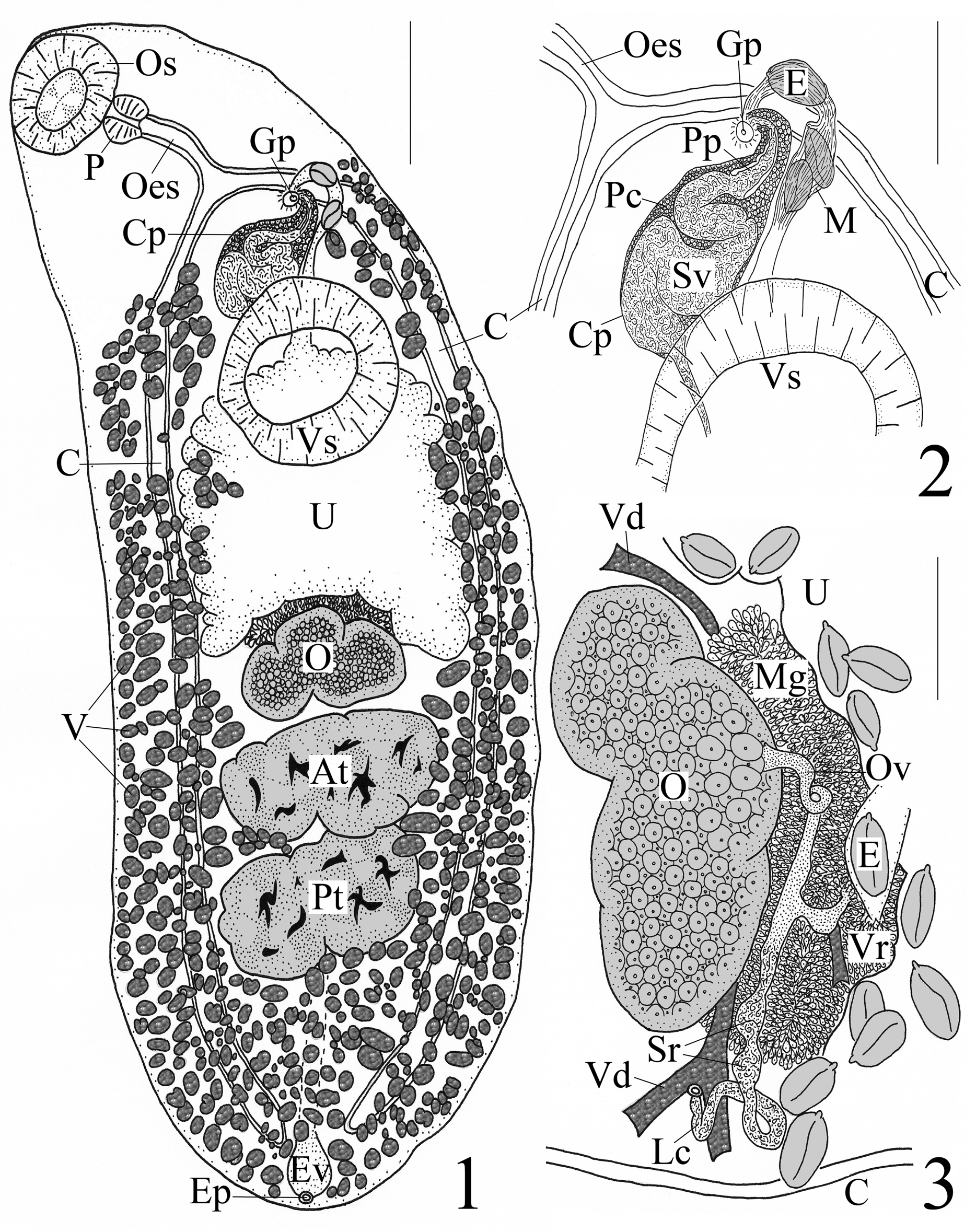
Re-description. [Based on 7 voucher specimens and 1 highly elongate and lanceolate specimen in {}. Measurements and proportions given in Table 3.] Body elongate-oval, 1 specimen highly elongate and lanceolate, widest post-equatorially; rounded at anterior and posterior extremities. Forebody attenuated anteriorly, narrows at bifurcal or oesophageal level. Hindbody rounded posteriorly and wider than forebody. Tegument smooth. Pre-oral lobe absent. Oral sucker subspherical, subterminal (terminal in 1 specimen). Ventral sucker sessile, spherical to subspherical, wider than long in most specimens, larger than oral sucker, post-bifurcal, pre-equatorial and located near junction of anterior and middle thirds of body. Prepharynx quite short, distinct. Pharynx muscular, spherical to dolioform. Oesophagus thick-walled, straight in most specimens [n=7] or slightly sinuous [n=1]. Intestinal bifurcation anterior to ventral sucker by (mean 191, range 40–272) [n=7] {336} long. Ceca narrow, run posteriorly along lateral sides of worm, end blindly near posterior extremity; cecal ends slightly arcuate and can be inflated.
Parasite T. beringi T. beringi T. beringi
n = 2 20 8
Host Coryphaenoides sp. Giant grenadier, Albatrossia Longfin grenadier,
( Macrouridae View in CoL ) pectoralis (Gilbert) View in CoL Coryphaenoides longifilis (Macrouridae) View in CoL Günther ( Macrouridae View in CoL ) ......continued on the next page Parasite T. beringi T. beringi T. beringi View in CoL 1 These specimens were collected by Dr. James M. McCauley (deceased), Oregon State University, and housed at the Harold W. Manter Laboratory (HWML), University of Nebraska, Lincoln, Nebraska, U.S.A., under these accession numbers. 2 These specimens were collected and described by TK as Tellervotrema katadara ( Kuramochi, 2001) View in CoL in Kuramochi (2009, pp. 30–31) and housed in the National Museum of Nature and Science, Tokyo, Japan (NSMT), under this accession number. 3 AT, anterior testis; L, length; PT, posterior testis; VS, ventral sucker; W, width; range with mean in parentheses; number [n] of measurements if different from total number of worms examined; one of eight specimens (NSMT-Pl 5672) appeared highly elongated and lanceolate in shape upon fixation (as opposed to elongate oval), and its measurements are included in {} to separate it from the seven others.
4 Proportion of body length.
Testes 2, tandem, smooth to lobed, transversely elongate, median, intercecal, post-equatorial mainly in posterior third of body, contiguous [n=3] or slightly separated [n=5]. Post-testicular region fairly sizeable, occupying posterior fourth of body. Cirrus pouch thin-walled, clavate, mostly pre-acetabular, extends anterosinistrally from midline to point about midway between midline and left margin, distinctly elongate in anterior portion with conspicuous sinistral turn over left cecum and toward left margin of worm in some individuals [n=4]; anterior extent of distal portion to level of intestinal bifurcation [n=6] or slightly posterior to bifurcation [n=2]; posterior extent of proximal portion either just overlaps anterior margin of ventral sucker [n=6] or extends to mid-level of latter [n=2]. Seminal vesicle bipartite; proximal portion saccular, convoluted; distal portion tubular, narrow, straight; terminates in small, circular, distinct genital atrium. Prostate gland-cells well developed, numerous in cirrus pouch with especially dense number in distal portion of pouch. Pars prostatica conspicuous, thick-walled, with numerous cells along walls, (67, 46–84) [n=7] {70} long × (27, 18–32) [n=7] {24} wide, in distal portion of cirrus pouch; ejaculatory duct distinct, straight, (66, 40–96) [n=5] {80} long. Genital pore submedian, sinistral, midway between left body margin and midline (located just left of midline in highly elongate/ lanceolate specimen), at level of intestinal bifurcation but can reach level of posterior oesophagus (post-bifurcal in highly elongate/lanceolate specimen), pre-acetabular by (311, 264–360) [n=7] {284} long, ventrally overlaps left margin of left cecum or short distance lateral or medial to left cecum, closer to ventral sucker than oral sucker.
Ovary moderate to deeply 3- to 4-lobed, compressed longitudinally in some individuals [n=4], median to dextrally submedian, intercecal, immediately pre-testicular, contiguous with anterior testis, post-equatorial in posterior portion of middle third of body. Oötype median, directly anterodorsal to ovary, anterior to anterodextral to vitelline reservoir. Mehlis’ gland cells conspicuous, numerous, overlapped by loops of uterus. Seminal receptacle canalicular, elaborate, overlaps left lobe of ovary and fills up large portion of Laurer’s canal via distinct swellings/ dilations in canal (cf. beads on a string) primarily anterosinistral to ovary. Laurer's canal with dorsal opening, (98, 68–128) [n=2] medial to left cecum (canal opening dorsally overlaps posterior margin of vitelline reservoir in highly elongate/lanceolate specimen), opening surrounded by dark-stained cells; canal itself extensive, convoluted, overlaps Mehlis’ gland-oötype complex and loops just before entering latter. Oviduct convoluted, arises from midline of ovary along its anterior margin. Vitellarium circumcecal, follicles numerous, circular or elongate oval or globular in shape, variable in size; anterior extent of vitellarium in forebody as 2 separate but at times indistinct “bunches” (i.e. Tellervotrema View in CoL vitellarium distribution) distributed from level of mid-oesophagus to level of posterior margin of ventral sucker, interruption in vitelline fields proximate to ventral sucker variable in number (0, 1 or 2 gaps), symmetry (gaps directly opposite ventral sucker, oblique to ventral sucker, or displaced longitudinally [highly elongate/lanceolate specimen with large gap in vitellarium in posterodextral region of worm]) and location (near body margin or displaced medially to near midline of worm towards and around ventral sucker); posterior extent of vitellarium in hindbody along lateral margins from level either near posterior margin of ventral sucker or between ventral sucker and ovary to posterior extremity, not confluent but encroaches over lateral margins of gonads, somewhat into immediate pre-ovarian region, space between ovary and anterior testis and into intertesticular area, completely confluent in post-testicular region [n=4] except over excretory vesicle [n=4]. Vitelline reservoir subtriangular, median to sinistrally submedian, indistinct in some individuals, directly dorsal to left lobe of ovary or anterodorsal to ovary or overlaps anterior margin of ovary. Vitelline ducts pass medially at level of anterior margin of ovary or just posterior to it. Uterus winding, intercecal, some loops pass ventrally over ceca but not lateral to them, fills up almost entire space between ovary and ventral sucker; wide proximal loops run anteriorly from ovary to posterior margin of ventral sucker then narrow and pass anterodorsal to either center or left margin of ventral sucker; distal loop turns anterosinistrally to run parallel along left side of cirrus pouch to terminate at genital pore. Metraterm indistinct, 142 [n=1] {70} long × 30 [n=1] {16} wide. Eggs smooth, oval, majority collapsed or crenulated, amber, non-filamented, operculate, with conspicuous knob or boss on one pole.
Excretory vesicle tubular, I-shaped, thin in width, extends to ovary along midline of worm. Excretory pore dorsal, subterminal, opens via a small sphincter; dark-stained cells surround pore and base of vesicle.
Remarks. The eight studied specimens conform to the diagnosis of the subfamily Plagioporinae and the genus Tellervotrema (macrourid host species = Coryphaenoides longifilis ) by Cribb (2005). Kuramochi (2009, p. 30–31) gave a rudimentary description of these same specimens (originally identified as T. katadara ) providing only a range of body lengths and widths (3.00–3.18 mm × 1.16–1.35 mm). He even stated that “… T. katadara may be closely related to T. beringi so far [as] measurements are concerned.”
This material compares well overall with the type description of T. beringi , originally described by Mamaev (1965) as Plagioporus beringi Mamaev, 1965 , from the stomach of seven specimens of an unidentified macrourid of the genus Coryphaenoides Gunnerus obtained at 150–500 m depths from 30 miles west of the Pribilof Islands, Alaska, and from near Cape Navarin, Russia, in the Bering Sea. When compared to the type description of T. beringi , we noted a few minor morphometric differences: the prepharynx of T. beringi in the original description was more distinct and longer (110 Μm vs 12–36 Μm); the pharynx had a slightly larger width (200 vs 112–144); the testes were larger (anterior testis: 360 × 780 vs 208–264 × 480–576; posterior testis: 380 × 750 vs 240–304 × 456– 608); the cirrus pouch had a slightly longer length (600 vs 388–476); and the ovary width was slightly smaller (340 vs 376–552). Blend et al. (2012) reported individuals of T. beringi in the intestine of the giant grenadier, Albatrossia pectoralis (Gilbert) , and Coryphaenoides sp. obtained from depths of 1,530 m and 2,800 m, respectively, in the northeastern Pacific Ocean off Oregon. The eight specimens of T. beringi re-described from C. longifilis here and in Table 3 have a slightly wider body, slightly wider testes and ovary, a longer distance between the ventral sucker and ovary, and larger vitelline follicles and uterus width; however, all other morphometric measurements for the present material along with the six proportions of body lengths (%) and two ratios given in Table 3 compare well to the specimens of T. beringi re-described by Blend et al. (2012).
The present material differs slightly from earlier descriptions of T. beringi in the posterior extent of the cirrus pouch relative to the ventral sucker and intestinal bifurcation. Mamaev (1965) reported the cirrus pouch of this species to reach to the posterior margin of the ventral sucker, and Blend et al. (2012) described it to at least overlap 1/5 to 3/4 the length of the ventral sucker and terminate well posterior to the level of the intestinal bifurcation. In these specimens, the posterior extent of the cirrus pouch just overlaps the anterior margin of the ventral sucker [n=6, 75%]—more like T. armstrongi— or extends to the mid-level of the ventral sucker [n=2; 25%]—like either T. armstrongi or T. beringi —and it terminates well posterior to the level of the intestinal bifurcation (see Blend et al. 2012).
Blend et al. (2012) commented that the testes texture of T. beringi from their material ranged from irregularly indented to deeply lobed; none were smooth as illustrated by Mamaev (1965, Fig. 1 View FIGURES 1 – 3 ) in the original description of this species. There were specimens in this study that possessed smooth testes, and we feel that this observation further supports the possibility of a high degree of plasticity in this feature within species of Tellervotrema ; an idea first hypothesized by Blend et al. (2012, Figs. 20–23), who on combining their observations of this feature with those of Mamaev (1965), concluded that testes texture of T. beringi can vary from smooth to irregularly indented to deeply lobed. Blend et al. (2012) also observed in T. beringi the characteristic vitellarium gap (separating the paired, isolated vitelline “bunches” in the forebody from the remainder of the vitellarium in the hindbody—this gap is characteristic of species of Tellervotrema ) at about the level of the posterior margin of the ventral sucker. However, the gap itself was quite variable in number (0, 1 or 2 gaps), symmetry (gaps directly opposite ventral sucker, oblique to ventral sucker, or displaced longitudinally to near level of gonads) and location (near body margin or displaced medially to near midline of worm) (see Blend et al. 2012, Figs. 11–16). This same variability in number, symmetry and location of the vitelline gap was seen in the present material of T. beringi from off Japan. Specifically, the gap in the vitelline fields proximate to the ventral sucker varied in number (0, 1 or 2 gaps), symmetry (gaps directly opposite ventral sucker, oblique to ventral sucker, or displaced longitudinally [the highly elongate/lanceolate specimen possessed a large gap in the vitellarium within the posterodextral region of the worm]) and location (near body margin or displaced medially to near midline of worm towards and around ventral sucker).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Plagioporinae |
|
Genus |
Tellervotrema beringi ( Mamaev, 1965 ) Gibson & Bray, 1982
| Blend, Charles K., Kuramochi, Toshiaki & Dronen, Norman O. 2015 |
Tellervotrema katadara (
| Kuramochi 2001 |