
Tapuiasaurus macedoi
|
publication ID |
https://doi.org/10.1111/zoj.12420 |
|
persistent identifier |
https://treatment.plazi.org/id/5B1D879C-FFA7-FFA6-A34D-58AAFEA8B115 |
|
treatment provided by |
Marcus |
|
scientific name |
Tapuiasaurus macedoi |
| status |
|
Below we discuss the evolutionary relationships of Tapuiasaurus macedoi based on a revised phylogenetic analysis of rescored character data from new observations and additional preparation. We then examine the role of missing data and the implications of the missing occurrences within particular strata in the original and revised results.
ZAHER ET AL. (2011) ANALYSIS
In their initial description of Tapuiasaurus macedoi, Zaher et al. (2011) performed a phylogenetic analysis that recovered Tapuiasaurus as a member of Nemegtosauridae , which also includes Rapetosaurus and Nemegtosaurus . This clade was positioned within Lithostrotia with moderate support, being the sister group of the clade formed by Isisaurus , Diamantinasaurus and Saltasauridae . Within Nemegtosauridae , Tapuiasaurus was hypothesized to be more closely related to Rapetosaurus than to Nemegtosaurus , a result that was considerably shorter than alternative arrangements, including one that placed it outside the two Late Cretaceous species (see Zaher et al., 2011: fig. 7).
Zaher et al. (2011) used a modified version of the Wilson (2002) matrix, which scored 27 terminal taxa for 234 characters. To this they added 12 cranial characters, some new and some from Curry Rogers (2005), as well as four terminal taxa ( Phuwiangosaurus , Tangvayosaurus , Diamantinasaurus and Tapuiasaurus ). Revised scorings for Euhelopus were used ( Wilson & Upchurch, 2009: table 6), as were previous scorings for Phuwiangosaurus and Tangvayosaurus ( Suteethorn et al., 2009) and Diamantinasaurus ( Hocknull et al., 2009) . Of the resultant 246 characters used by Zaher et al. (2011), there are 88 cranial characters (35.8%), 72 axial characters (29.3%), 85 appendicular characters (34.6%) and one dermal character (0.4%). Of the 31 resulting terminal taxa, 12 (39%) are titanosaurs.
The taxonomic scope of the Wilson (2002) matrix was Sauropoda, which ranges from the Late Triassic to the latest Cretaceous. Zaher et al. (2011) repurposed that matrix to focus on a much narrower taxonomic scope, Titanosauria, which is restricted to the Cretaceous ( D’Emic, 2012). Wilson (2002) included eight titanosaurs in his analysis, and although this represents 30% of the terminal taxa, it accounts for only a small percentage of the 70+ species currently recognized to comprise that clade (J. A. Wilson & M. D. D’Emic, unpubl. data). Although there was good character support for two of the seven nodes within Titanosauria (decay index = 4) and moderate support for another (decay index = 2), three nodes had a decay index of 1 ( Wilson, 2002: table 12). With the addition of four more titanosaur terminal taxa in the Zaher et al. (2011) analysis, the original character budget was stretched across 50% more nodes. Even with the addition of 12 new characters by Zaher et al. (2011), we might expect reduced levels of Tapuiasaurus (all characters; 1 – 246)
00?1011201 0110010101 0 – 02011111 0111121111 0?10001000 010110100??00 – 111101 121000110? 1111011?0 –?1010?1?10 1100110??? ?????????? ?????????? ????????10 11??????1? ????0?0??? ????110010 1????????? ?????????? ?????1?2?? ?????????? ??????1??? ?????0?11? 1110110111 111111
Isisaurus (cranial characters only; 1 – 88)
?????????? ????????0? 0?0?0????? ?????????? ??1?0??000 0??1?????? ?????????? ?????????? ????????
The only scorings that changed between the two analyses are those of Isisaurus and Tapuiasaurus , which are labelled ‘orig’ (original) or ‘rev’ (revised) accordingly. Missing data have been broken down and ranked for total missing data, cranial missing data and postcranial missing data. Taxa are listed in order of their ‘Rank Sum’, which indicates the sum of the rank scores for total, cranial and postcranial missing data. Note that each of the four rankings provides a different ordering of taxa.
support within Titanosauria (see Whitlock, D’Emic & Wilson, 2011 for discussion of ‘diluent’ taxa).
The phylogenetic position of Tapuiasaurus within a clade formed by the latest Cretaceous Rapetosaurus and Nemegtosaurus was not robustly supported (decay index = 1) by the data assembled by Zaher et al. (2011). Other relationships within Titanosauria had better support, with decay indices of 2 and 3. Phylogenetic tests using constraint trees demonstrated that the published topology was significantly shorter than alternative arrangements placing Tapuiasaurus in a more basally diverging position.
RE- ANALYSIS
Owing to the low level of support for the monophyly of Nemegtosauridae in the original analysis, combined with broad taxonomic scope of the Wilson (2002) matrix, extensive missing data and lengthy implied ghost lineages (see below), we direct the new morphological data described here for Tapuiasaurus towards a re-analysis of its phylogenetic position.
Our modifications to the Zaher et al. (2011) matrix were restricted to rescoring the cranial and postcranial data for Tapuiasaurus and scoring cranial data for Isisaurus . No other matrix cells were changed ( Table 6). The revised Tapuiasaurus scoring contains substantially fewer missing entries than the original analysis ( Table 7). Most of the disambigua- tions (i.e. replacing a ‘?’ with a positive score) were localized within the skull, for which Tapuiasaurus now has the lowest missing data score for any terminal taxon (4.5% missing cranial data). Although there were several disambiguations in other parts of the skeleton, Tapuiasaurus still remains very incompletely scored postcranially, second only to Nemegtosaurus (100% incomplete) in postcranial missing data.
In addition to the new scorings for Tapuiasaurus , we added scorings for a braincase and skull roof of Isisaurus , based on relatively new links between these elements and the holotypic postcranial skeleton ( Wilson et al., 2005, 2009). Only ten additional data cells were filled for Isisaurus .
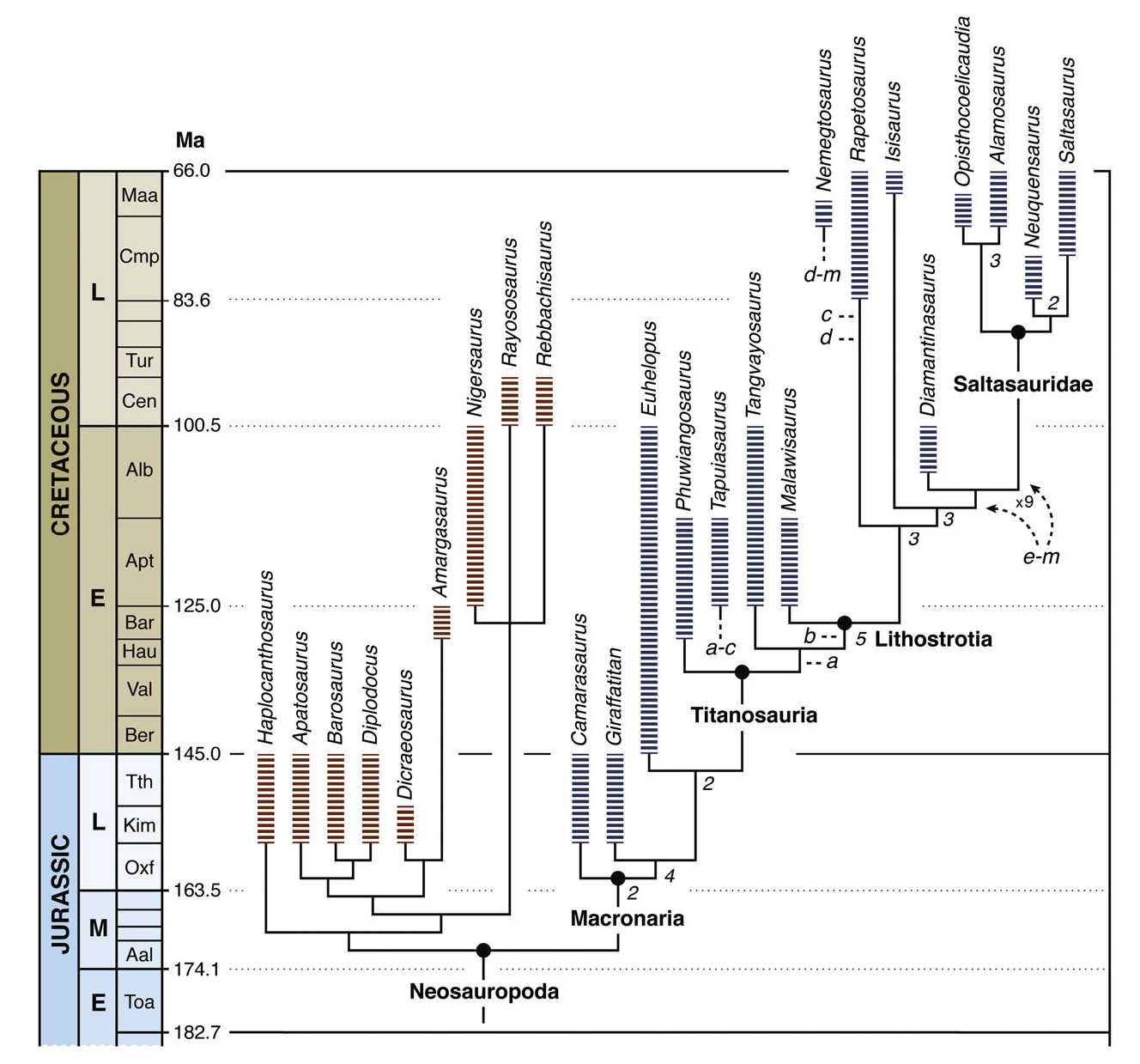
The rescored phylogenetic dataset of 27 terminal taxa for 246 characters was analysed under equally weighted parsimony using TNT v.1.1 ( Goloboff, Farris & Nixon, 2008a,b). A traditional heuristic tree search was conducted in which 1000 replicates of Wagner trees were created using random addition sequences of taxa, followed by tree bisection-reconnection (TBR) branch swapping. A final round of TBR was applied to the most parsimonious trees (MPTs) found in the replicates. Thirty-four MPTs were found after this heuristic tree search of 462 steps (CI = 0.593, RI = 0.770). The strict consensus of these 34 MPTs shows a large polytomy involving all titanosaurs. Evaluation of the topological variation among the 34 MPTs using iterPCR ( Pol & Escapa, 2009) identified Nemegtosaurus and Tapuiasaurus as the two unstable taxa that caused the large polytomy among titanosaurs. A reduced strict consensus showing the six alternative positions of Nemegtosaurus and the three alternative positions of Tapuiasaurus within an other- wise completely resolved topology for Titanosauria is shown in Fig. 23 View Figure 23 .
In contrast to the original analysis, the rescored analysis does not unequivocally resolve Tapuiasaurus , Rapetosaurus and Nemegtosaurus as a monophyletic group. Although this topology is retrieved in two of the 34 MPTs ( Fig. 23 View Figure 23 , letters c and d), all other most parsimonious topologies depict Tapuiasaurus more basally than in the original analysis: as the sister group of either Lithostrotia or Tangvayosaurus + Lithostrotia ( Fig. 23 View Figure 23 , letters a and b). The alternative positions of Nemegtosaurus , in contrast, are within or adjacent to Saltasauridae ( Fig. 23 View Figure 23 , letters e – m). Rapetosaurus is placed in an equivalent position to that of the original analysis (within Lithostrotia and basal to Isisaurus and saltasaurids). The affinities of Tapuiasaurus with Lithostrotia are based on character data present in the original analysis, such as the posterolaterally orientated quadrate fossa (char. 35.1), basisphenoid – quadrate contact (char. 52.1) and reduced cervical neural arch lamination (char. 81.1). The key difference with respect to the previous result is that Tapuiasaurus is placed in some of the MPTs outside Lithostrotia, a basal position supported in those trees by the absence of derived characters shared by Malawisaurus and/or Rapetosaurus and more derived titanosaurs: presence of osteoderms (char. 234.1), simple undivided cervical pneumatopores (char. 83.0), mid-posterior dorsal neural spines orientated posteriorly (char. 104.1), cylindrical tooth crowns (char. 70.2), coracoid proximodistal length twice that of the scapular articulation (char. 155.1) and distal radius breadth about twice that of the radial midshaft (char. 170.1).
The support values for most nodes within Titanosauria are extremely low (e.g. decay index = 0, bootstrap/jackknife frequencies below 50%). If we ignore the alternative positions of the unstable Nemegtosaurus and Tapuiasaurus among suboptimal trees (for decay index) or trees found in the bootstrap/ jackknife pseudoreplicates, then support values are markedly higher for basal nodes of Titanosauria (decay indices = 3 – 5, bootstrap/jackknife frequencies = 63 – 80%). This indicates two important facts. First, the phylogenetic position of two taxa known primarily from skull anatomy ( Tapuiasaurus and Nemegtosaurus ) must be regarded as highly labile. Second, the addition of new information on Tapuiasaurus and Isisaurus reveals character conflict, previously hidden by missing data, that makes Tapuiasaurus and Nemegtosaurus unstable in the revised dataset.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.