
Sasanidus, Freyhof, Jörg, Geiger, Matthias F., Golzarianpour, Kiavash & Patimar, Rahman, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4107.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:21FB8099-278A-4F44-B207-717683A0D5B0 |
|
DOI |
https://doi.org/10.5281/zenodo.6057150 |
|
persistent identifier |
https://treatment.plazi.org/id/59377E3D-B76B-FFF6-FF7C-FADBFA6DFDFE |
|
treatment provided by |
Plazi |
|
scientific name |
Sasanidus |
| status |
gen. nov. |
Sasanidus , new genus
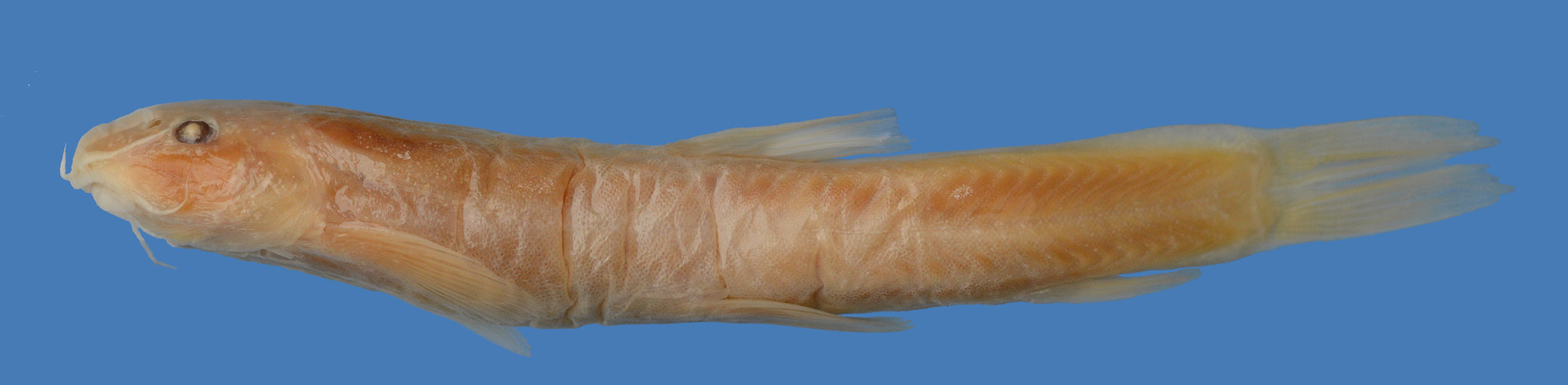
Fig. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4
Type species. Noemacheilus kermanshahensis Bănărescu & Nalbant, 1966:151 .
Material examined. All from Iran: FSJF 3213, 5, 46–63 mm SL; Hamadan prov.: Gamasiab River, 34°13'N 48°17'E. — FSJF 3234, 5, 30–45 mm SL; Kermanshah prov.: Sepidbarg River west of Javanrud, 34°48'21.44"N 46°27'29.15"E. — FCKG 802, 21, 35–45 mm SL, Lorestan prov.: stream below Horro bridge at Dorood- Khoramabad highway, 33°31'13.8"N 48°46'19.0"E. — FCKG 806, 3, 45–70 mm SL, Lorestan prov.: stream Ab- Alashtar on road from Alashtar to Khoramabad, 33°47'09.6"N 48°12'23.0"E. — FCKG 805, 4, 38–47 mm SL, Lorestan prov.: Sezar (Marboreh) River at Dorood, 33°28'38.9"N 49°03'59"E. — FCKG 800, 40, 38–71 mm SL; Hamadan prov.: spring Pashow near Gian, flowing to Gamasiab River, 34°13'N 48°17'E. — FCKG 804, 1, 70 mm SL; Hamadan prov.: Gamasiab River at Baba-Rostam, 34°07'55.7"N 48°23'55.1"E. — FCKG 801, 21, 32–45 mm SL; Hamadan prov.: Haram-Abad River at Ashmizan, 34°07'N 48°85' E. — FCKG 807, 20, 32–45 mm SL; Hamadan prov.: stream seyyed Shahab, Bujin village, Asad-abad, 34°48'10.9"N 48°02'42"E. — FCKG 803, 2, 35–40 mm SL; Lorestan prov.: stream Kakareza at Kakareza village, 33°42'35.9" N 48°16'38.7"E. — FCKG not catalogued, 6, 35- 45 mm SL; Lorestan prov.: Houzian river, road Aligudarz toward Gahar lake, 33°21'31"N 49°44'24"E.
Material used in the molecular genetic analysis. FSJF DNA-1997; Iran: Kermanshah prov.: Sepidbarg River west of Javanrud, 34°48'21.44"N 46°27'29.15"E (GenBank accession numbers: KU928261 View Materials , KU928288 View Materials , KU928289 View Materials ).
Diagnosis. Sasanidus is not diagnosed by any unique external character state, but by a combination of characters. Sasanidus was earlier placed in the genus Oxynoemacheilus and is distinguished from all species in that genus and also from all species of Seminemacheilus by the absence of an external sexual dimorphism (vs. longer pectoral fin, nuptial tubercles on fins, head and back in male). The prominent, irregularly shaped midlateral stripe, being characteristic for S. kermanshahensis , is rarely found in species of Oxynoemacheilus , but present in a few species ( O. angorae , O. phoxinoides ). Not all individuals of S. kermanshahensis show the characteristic midlateral stripe, which is often restricted to the caudal peduncle, and some have a mottled or marbled colour pattern ( Fig. 2 View FIGURE 2 ). Sasanidus is also distinguished from Seminemacheilus by having one epural bone in the caudal skeleton (vs.
absent). Sasanidus is distinguished from Paracobitis by the absence of a dorsal adipose crest (vs. presence), the absence of a dark brown or black bar or blotch at the caudal-fin base (vs. presence) and the presence of a prominent whitish or yellowish triangular patch on the posteriormost upper and lower caudal peduncle (vs. absence). Sasanidus is distinguished from Turcinoemacheilus by having the pelvic-fin origin clearly behind a vertical of the dorsal-fin origin (vs. pelvic-fin origin in front of a vertical of the dorsal-fin origin) and the anus situated about one eye diameter in front of the anal-fin origin (vs. more than two eye diameters). Sasanidus is distinguished from Paraschistura by lacking the black jet spot at the dorsal-fin origin (vs. presence), having a marbled colour pattern with a prominent, irregularly shaped midlateral stripe (vs. lateral bars often dissociated into irregularly shaped blotches or faded on flank in front of the dorsal-fin origin) and a slightly emarginate caudal fin (vs. deeply emarginate or forked), no dark brown or black bar or blotch at caudal-fin base (vs. presence) and prominent whitish or yellowish triangular patch on the posteriormost upper and lower caudal peduncle (vs. absence).
Etymology. The genus is named for the Sassanid Empire, which was recognized as one of the leading regional powers for a period of more than 400 years. During Late Antiquity, the Sasanian Empire is considered to have been one of Iran's most important and influential historical periods. Gender masculine.
Distribution. Sasanidus kermanshahensis is known from headwaters of the Karkheh and Dez Rivers. The Dez is a tributary to the Karun and both the Karkheh and Karun flowi from the Iranian Zagros Mountains westwards into the lowermost Tigris ( Abdoli 2000).
Included species. Monotypic.
Discussion. The COI barcode sequences analysed here and the very limited species coverage of the speciesrich family Nemacheilidae do not allow strong phylogenetic conclusions to be made from the dataset presented in this study. Therefore, we do not speculate on the relationships of the different genera of nemacheilid loaches included in this study. The holotype of Sasanidus kermanshahensis was illustrated by Bănărescu & Nalbant (1966:152) and the species can be immediately identified by its body shape and colour pattern. The osteology of Sasanidus has recently been described by Mafakheri et al. (2015) and osteological characters mentioned below were examined by us based on our own materials. Sasanidus belong to a group of nemacheilid genera distinguished by the absence of the preethmoid I in the ethmoid region of the skull. Other genera in this group are Acanthocobitis , Afronemacheilus , Dzihunia , Indoreonectes , Iskandaria , Mesonoemacheilus , Nemacheilus , Oxynoemacheilus , Paracobitis , Paraschistura , Schistura , Seminemacheilus and Troglocobitis ( Prokofiev 2009) . It remains unclear, if this group of genera forms a monophyletic group of related species. Afronemacheilus occurs in East Africa, Troglocobitis , Dzihunia and Iskandaria are found in the Aral Sea basin in Central Asia. Sasanidus is distinguished from Afronemacheilus by having a single epural bone in the caudal skeleton (vs. absent). It is distinguished from Troglocobitis by having a bony swim-bladder capsule (vs. absent) and from Dzihunia by having simple lip structures (vs. lower lip with pillow-like elements interrupted by deep folds) and a smooth skin (vs. with small, spine-like epidermal tubercles). Sasanidus is distinguished from Iskandaria by having the body fully covered by scales (vs. no scales), a deep body and caudal peduncle (vs. very slender and shallow) and no muscular, swollen cheeks in males (vs. present).
There are few studies about the genera of nemacheilid loaches of the Middle East ( Bănărescu & Nalbant 1995; Kottelat 1990; Prokofiev 2009) and there are also few studies presenting molecular data on nemacheilid loaches of that area (Esmaeili et al. 2014; Freyhof et al. 2014; Freyhof et al. 2015). There is no comprehensive molecular study on the nemacheilid loaches of India or South-East Asia. Here, we bring all known genera of nemacheilid loaches from the Middle East together in one molecular dataset for the first time. Furthermore, we were able to add COI sequence data from representatives of all genera of nemacheilid loaches recognised as valid from western India and Pakistan by Kottelat (2012).
However, a phylogenetic study cannot be based solely on the molecular method applied here, because the resolution of this single mitochondrial marker is insufficient to reliably resolve the relationships between the different genera studied. DNA barcode data do not offer a phylogenetic signal sufficiently strong to resolve evolutionary relationships at higher taxonomic levels ( Hajibabaei et al. 2006), which is clearly reflected in the low bootstrap support values of the inner nodes ( Fig. 1 View FIGURE 1 ). Although analysed by phylogenetic-tree reconstruction methods, barcode-based trees should therefore not be interpreted as phylogenetic trees ( Hajibabaei et al. 2007). The method is well-suited to assign specimens to species, supported by high bootstrap values of nodes towards the tips, and mark conspicuous populations or taxa. In our study, a ML tree helped to identify Sasanidus as candidate taxon for a more comprehensive investigation, and aided with taxonomic sampling, putting the mitochondrial lineage of these fishes into a broader phylogenetic and biogeographic context. Taking all concerns into consideration, our data does not even place Sasanidus close to one of the genera described from the Middle East or India, supporting the hypothesis that it represents a distinct genus of its own. However, we do not exclude that other species of Sasanidus might occur in Pakistan or India, where they might currently be identified as species of Schistura . Nevertheless, none of the species described by Mirza et al. (1981) from Pakistan and by Menon (1987) from India are even superficially similar to Sasanidus .
It is well known that Schistura is a polyphyletic assemblage of species ( Bănărescu & Nalbant 1995; Kottelat 1990; Prokofiev 2009; Kottelat 2012). To exclude the possibility that Sasanidus is a member of Schistura , we examined two syntypes of S. rupecula , the type species of Schistura . Schistura rupecula was described from the city of Shimla in Himachal Pradesh, India ( McClelland 1838). The stream in Shimla is highly polluted today and no loaches could be found (Nikhil Sood, Bangalore; Rajeev Raghavan, Cochin, pers. comm.). The stream flowing through Shimla is a headwater stream of the Aasan River, which flows to the Giri River, itself being a tributary to the Ganges. We include here one DNA sequence of a loach from the lowermost Aasan River from our own material, just at the confluence with the Giri River. We compared the series (FSJF 3514; Fig. 4 View FIGURE 4 ) to which this individual belongs, with the description and figure by McClelland (1838:948), two syntypes of S. rupecula (SMF 8993; Fig. 6 View FIGURE 6 ) and an illustration of a possible syntype of S. montana (BMNH 1860.3.19.118), a second species described from Shimla by McClelland (1838:948). All these loaches have 5½ branched anal-fin rays, 8+8 branched caudal-fin rays, bars on the flank and a moderately deep emarginate caudal fin. Despite being geographically very close to the type locality of S. rupecula and S. montana , the fishes from Aasan River could not definitively be identified as one of these two species and are referred to as Schistura sp. here.
In addition to one DNA sequence of Schistura sp. from the Aasan River, we included the COI sequence data of three Schistura spp. From the same area and 26 sequences of fishes identified as belonging to the genus ‘ Schistura ’ from NCBI GenBank ( Table 1). We sequenced four additional ‘ Schistura ’ semiarmata and 11 individuals of four unidentified species of ‘ Schistura ’ from the Western Ghats that form a separate evolutionary lineage ( Fig. 7 View FIGURE 7 ). The various loaches tentatively placed in the genus Schistura and analysed in this study are not closely related and their clustering implies that several undescribed genera might be involved. Importantly, none of the mitochondrial lineages designated as ‘ Schistura ’ in Figure 7 View FIGURE 7 are closely related to Sasanidus . Future studies on the diversity of loaches in Pakistan and India are encouraged to resolve their generic structure and the biogeographic relationships.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |