
Plesiadapis
|
publication ID |
https://doi.org/ 10.1206/3797.1 |
|
persistent identifier |
https://treatment.plazi.org/id/587387C9-1431-517F-FE18-1E13FCA4FE9E |
|
treatment provided by |
Carolina |
|
scientific name |
Plesiadapis |
| status |
|
Plesiadapis cf. P. churchilli Lofgren et al., 2008
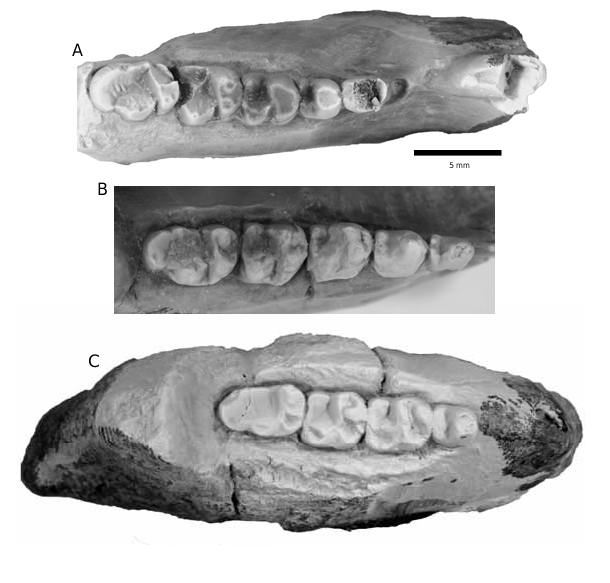
HOLOTYPE: RAM 9044 , right dentary fragment with damaged i1, p2 alveolus, and p3–m3 from RAM locality V200510, member 4a, Goler Formation , California ( fig. 3A View FIGURE 3 ).
REFERRED SPECIMENS: RAM 9668, left maxilla fragment with C, alveoli of P2 and P3, P4–M1 ( fig. 5C View FIGURE 5 ), and RAM 9669, crushed skull fragment with broken left I1, right P4, M1–2 and associated right p3 ( fig. 5E–F View FIGURE 5 ) from RAM locality V200510, member 4a; RAM 9429, right dentary fragment with i1 root and p3–m3 from RAM locality V200706, member 4a ( fig. 3B View FIGURE 3 ); RAM 10029, right dentary fragment with damaged p4–m3 from RAM locality V200804, member 4a ( fig. 3C View FIGURE 3 ); five m3s RAM 6961 (fig. 4A), RAM 6963 (fig. 4B), RAM 6960 (fig. 4C), RAM 6700 (fig. 4D), RAM 6934, m2 RAM 6446, mx fragment RAM 6935, mx trigonid RAM 6946, M3 RAM 6957 ( fig. 5H View FIGURE 5 ), M2 RAM 6962 ( fig. 5G View FIGURE 5 ), P3 RAM 6931 ( fig. 5D View FIGURE 5 ), and 2 I1s RAM 6431 ( fig. 5A View FIGURE 5 ), RAM 6929 ( fig. 5B View FIGURE 5 ) from RAM locality V98012 View Materials , member 4b; RAM 7193, partial right dentary with talonid fragment of p4, m1 and broken m2 from RAM locality V200202, member 4b; RAM 6925, partial left m3 from RAM locality V94014 View Materials , member 4a.
ETYMOLOGY: Named in honor of Richard “Dick” Lynas, for his dedicated efforts in support of collection and study of Goler Formation vertebrate fossils.
DIAGNOSIS: Midsized plesiadapid with a narrow p4 and m2 that align it with Nannodectes and a unique combination of other features that separates it from all known plesiadapid species, which include, I1 with mediocone small or absent and centroconule absent, upper molars with no significant development of crenulated enamel and lacking mesostyles, p3–4 with inflated protoconids and distinct talonid basins, m1–2 with squared entoconids, and m3 with slight to moderate talonid enamel crenulations and broad and unfissured hypoconulid that is usually rounded.
DESCRIPTION: The holotype of Nannodectes lynasi (RAM 9044) has a damaged i1, p3, m2, and m3 ( fig. 3A View FIGURE 3 ). Only the triangular-shaped posterior part of i1 is preserved. A single round depression, posterior to i1 and directly anterior to p3, represents the p2 alveolus. The trigonid of p3 is missing due to breakage. The talonid of p3 has two small medial cusps on its posterior margin (labially positioned cusp largest), and a very small, but distinct basin is present. The trigonid of p4 is dominated by a massive and worn protoconid (if a paraconid and metaconid were present, all trace has been removed by wear), and a distinct cristid obliqua connects the trigonid to a large hypoconid. The posterior margin of the talonid is defined by a postcristid developed between the hypoconid and smaller entoconid. The talonid basin of p4 is large, but relatively small compared to m1–
m2. Molar trigonids are simple,
with large metaconids and protoconids, and smaller labially shifted paraconids (breakage removed metaconid and paraconid of m3).
Talonids of m1–2 have large hypoconids and the entoconid of m1 is squared (entoconid of m2 broken).
Crenulated enamel is not developed on m1–2. Notable features of the m3 talonid are faint to moderately developed crenulations, a
FIGURE 4. Occlusal views of m3s of Nannodectes lynasi from RAM labially projecting hypoconid lobe, locality V98012 View Materials . A. RAM 6961, right m3. B. RAM 6963, left m3. and a broad, rounded, and unfis- C. RAM 6960, right m3. D. RAM 6700, right m3. sured hypoconulid. RAM 9429 represents another mostly complete dentary of N. lynasi ( fig. 3B View FIGURE 3 ), but one with significant enamel corrosion. RAM 9429 has a small, laterally crushed depression directly anterior to p3 that may represent the p2 alveolus, and a large, anteroposteriorly elongate protoconid dominate the trigonid of p3. The entoconid, hypoconid, cristid obliqua, and talonid basin are more developed in the p3 of RAM 9429 than the p3 of the holotype (RAM 9044). The trigonid of p4 has a very large protoconid, but unlike the holotype, a small metaconid and labially shifted paraconid are also present. Similar to p3, the hypoconid and entoconid are larger, and cristid obliqua and talonid basin more developed in the p4 of RAM 9429 than the holotype. The m1 of RAM 9429 has a large protoconid and metaconid and a smaller, lingually positioned paraconid (less lingually positioned than in p4). In m2, the paraconid is more lingually positioned and more closely appressed to the metaconid. Entoconids of m1 and m2 are squared in RAM 9429. The talonid of m3 has a broad, rounded, and unfissured hypoconulid like the holoype, but differs in having a narrower hypoconulid lobe and a small entoconid that forms a distinct lingual bulge. RAM 10029 is a partial dentary with heavily damaged p4–m3 as the crowns are partially dissolved ( fig. 3C View FIGURE 3 ). Only the most robust crown features of RAM 10029 are visible, but in general shape and size, RAM 10029 closely resembles the holotype (RAM 9044) and RAM 9429. Also, the p4 talonid basin of RAM 10029 was large, like the p4 of RAM 9429. RAM 10029 also has a broad and rounded m3 hypoconulid and lacks an entoconid lobe, features more like the holotype than RAM 9429. The partial dentary (RAM 7193) and isolated m3s previously referred to Plesiadapis cf. P. churchilli or P. cf. P. anceps ( Lofgren et al., 2008) , compare favorably to the holotype of Nannodectes lynasi and RAM 9429, except for RAM 6963. Isolated m3s of N. lynasi from the Goler Formation are uniform in morphology (fig. 4A, C–D), but RAM 6963 is smaller than all other m3s ( table 3) and its hypoconulid is distinctly squared (fig. 4B). Because RAM 6963 comes from the same site as four other m3s of N. lynasi (fig. 4A, C–D), it most likely represents an extreme morphological variant of the species.
Upper incisors of N. lynasi are broken, but RAM 6929 and RAM 6431 preserve a significant part of the crown ( fig. 5A–B View FIGURE 5 ), and RAM 9669 just the alveolar rim. RAM 6929 and RAM 6431 were described by Lofgren et al. (2008), and their important features are the minute mediocone and lack of a centroconule in RAM 6929 and the absence of both a mediocone and centroconule in RAM 6431 ( fig. 5A–B View FIGURE 5 ). The alveolar rim of RAM 9669 is 2.54 mm in width and 4.34 mm in length.
The upper canine and premolars of N. lynasi are represented by RAM 9668 (C and P4, with alveoli of P2–3), RAM 6931 (P3), and RAM 9669 (P4). In RAM 9668 ( fig. 5C View FIGURE 5 ), a single-rooted tooth is positioned adjacent to the anterior alveolus of P2, indicating the lack of a diastema between the teeth. This rarely preserved tooth has been identified as the canine in plesiadapids ( Gingerich, 1976; Boyer, 2009), although at first glance it would appear to be P1 based on its
A B
position adjacent to P2. The canine of RAM 9668 is a single upright cusp elongated anteroposteriorly, with a small shelf positioned low on the anterior face of the tooth. The close placement of the canine and P 2 in RAM 9668 is reminiscent of the condition seen in Wa 282, a partial maxilla of Plesiadapis walbeckensis . In contrast, a canine to P2 diastema is present in PU13393, a partial maxilla of P. fodinatus . Alveoli present anterior to the P 4 in RAM 9668 indicate that P2 had two circular roots orientated anteroposteriorly, with the anterior one approximately half the diameter of the posterior root and P3 had two roots of equal size, oriented transversely. Diastemas apparently were not developed between premolars in RAM 9668.
The only known P3 of Nannodectes lynasi (RAM 6931), described by Lofgren et al. (2008) as a P4, lacks a metacone and metaconule, but has a prominent protocone, and a paracone with a paraconule developed on its lingual face ( fig. 5D View FIGURE 5 ). Both P4s (RAM 9669 and RAM 9668) of N. lynasi have a closely appressed paracone and metacone, small parastyle, prominent paraconule with a small enamel bulge positioned posteriorly (that could represent an incipient metaconule), large protocone, well-developed anterior and posterior cingulum, and weakly developed stylar shelf and labial cingulum ( fig. 5C, E View FIGURE 5 ). Also, the lingual cingulum is not continuous in P4 and the mesostyle and metastyle are absent. In contrast to P3, whose paraconule is positioned on the lingual face of the paracone, the paraconule and paracone of P4 are separated by a distinct valley.
Features of the M1 and M 2 in N. lynasi are similar and include: widely spaced paracone and metacone subequal in size and connected by centrocrista, metacone positioned posteriorly, parastyle small and connected to paracone by weak paracrista, ectoflexus weak, labial cingulum present, protocone larger than metacone and paracone and connected to paraconule and metaconule by preprotocrista and postprotocrista, small hypocone present on crest extending posteriorly from protocone that merges with postcingulum, well-developed precingulum, lingual cingulum not continuous across protocone, and mesostyle and metastyle absent ( fig. 5C, F, G View FIGURE 5 ). The only differences between M1 and M2 are a more developed centrocrista and parastyle on M2, and a stronger M2 ectoflexus in RAM 6962 ( fig. 5G View FIGURE 5 ), compared to both the M1 of RAM 9668 and the M1–2 of RAM 9669 ( fig. 5C, F View FIGURE 5 ). RAM 6957, the only known M3 of N. lynasi ( fig. 5H View FIGURE 5 ) is damaged and has slightly crenulated enamel and lacks a distinct hypocone ( Lofgren et al., 2008). Measurements of the dentition of N. lynasi are provided in tables 3–4.
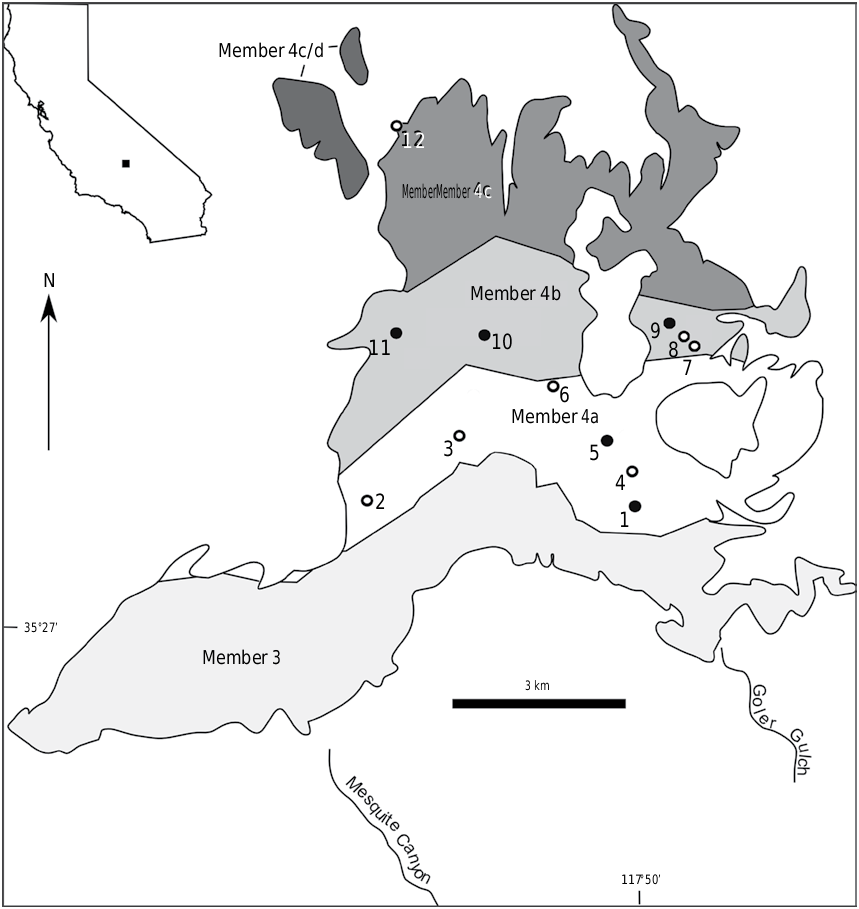
DISCUSSION AND COMPARISONS: The initial sample of this Goler plesiadapid consisted of 14 isolated teeth and a dentary fragment found before 2003 that were referred to Plesiadapis cf. P. churchilli (dentary fragment) and P. cf. P. anceps (isolated teeth) because the specimens could not be confidently assigned to a known species (Lofgren at al., 2008). Between 2007 and 2009, three partial dentaries and two partial maxillae were recovered from a 300 m thick sequence of member 4a strata in a large canyon informally termed Grand Canyon (no. 2 in fig. 1 View FIGURE 1 ). RAM locality V 200510 in Grand Canyon yielded a dentary with p3–m3 (RAM 9044) and two partial upper dentitions (RAM 9668, RAM 9669), specimens reasonably certain to represent the upper and lower dentition of a single species.
During the early and middle Tiffanian in North America, lineages of similar plesiadapid genera, Nannodectes and Plesiadapis , evolved in parallel, with Nannodectes distinctly smaller than Plesiadapis when both genera are found at the same site ( Gingerich, 1976). Also, Nannodectes was distinguished from contemporaneous species of Plesiadapis by its narrower cheek teeth (especially in the lower premolars), more triangular P3 (usually lacking a paraconule), and lack of a centroconule on I1 ( Gingerich, 1976).
In the Goler Formation, a single plesiadapid species is present, so size comparison between morphotypes is not possible. Also, the lack of a centroconule on I1 and a more triangular P3 are characters that do not reliably distinguish the two genera as they are found in Nannodectes and some species of Plesiadapis . However, narrower cheek teeth appear to distinguish Nannodectes and Plesiadapis as a cladogram based on dental characters of plesiadapid species indicates that Nannodectes is monophyletic, based on proportional differences between length and width of p4, M1–3, and m1–3 ( Boyer, 2009: fig. 5.1 View FIGURE 5 , table 5.1, characters 18, 23, and 31). Thus, the lower cheek teeth of Nannodectes should be narrower than those of Plesiadapis and upper molars of Nannodectes more squared compared to more rectangular upper molars of Plesiadapis . To test this, ratios of length versus width of cheek teeth of Nannodectes and North American species of Plesiadapis were calculated using length and width means provided by Gingerich (1976: table 2, tables A3–A 13) for each species. These ratios were compared to similarly derived ratios for the cheek teeth of Nannodectes lynasi from the Goler Formation ( table 5). For the lower dentition, a number above 1.00 for a tooth site indicates that the teeth are longer than wide (narrower). For the upper dentition, all teeth are wider than long, but a number close to 1.00 for a tooth site indicates that these teeth are more squared (less rectangular).
Length versus width ratios of cheek teeth provide limited support for distinguishing Nannodectes from Plesiadapis . Ratios of the p4 of N. intermedius , N. gazini , N. simpsoni , and N. gidleyi are all above 1.00, indicating that the p4 of Nannodectes is consistently narrow. Early and middle Tiffanian species of Plesiadapis ( P. praecursor , P. anceps , P. rex , P. churchilli ) have a p4 that is wider than long (ratio less than 1.00), but later Tiffanian species ( P. fodinatus , P. dubius , P.cookei ) have narrow p4s like Nannodectes ( table 5). The m2 of Nannodectes is slightly narrower than the m2 of Plesiadapis praecursor , P. anceps , and P. rex , but not narrower than the m2 of P. churchilli , P. fodinatus , P. dubius , and P. cookei ( table 5). For the p3, m1 or m3, there is no significant difference in length versus width ratios between genera. For the upper dentition, the P3 of Nannodectes gazini , N. simpsoni , and N. gidleyi is more squared than in any North American species of Plesiadapis , while the P4 of Nannodectes gazini , N. simpsoni , and N. gidleyi is significantly more squared than only Plesiadapis praecursor , P. rex , and P. churchilli ( table 5). There are no significant differences in ratios of length and width in the upper molars of Nannodectes and North American species of Plesiadapis ( table 5). Length versus width ratios indicates that only a narrow p4 and more squared P3 and P4 are measurable characters that separate Nannodectes from early and middle Tiffanian species of Plesiadapis .
Goler dentaries (RAM 9044, RAM 9429, and RAM 10029) have narrow p4s and m2s with ratios above 1.00 ( table 5), which supports their referral to Nannodectes , but Goler P4s and the single P3 have ratios not clearly aligned with either Nannodectes or Plesiadapis ( table 5). Thus, proportional differences in certain cheek teeth of Nannodectes and Plesiadapis that distinguish the two genera are valid only for the lower dentition of the Goler sample. However, comparison of the dentition of this Goler plesiadapid to all species of Nannodectes , Plesiadapis , and Chiromyoides indicates a closer morphological affinity to Nannodectes gidleyi and N. aff. gidleyi , which also supports referral of the Goler sample to Nannodectes and the diagnosis of the new species, N. lynasi .
When compared to middle and late Tiffanian species of Plesiadapis , Nannodectes lynasi is much smaller than Plesiadapis simonsi , P. gingerichi , P. cookei , P. tricuspidens , and P. remensis , and lacks the more derived dental features seen in some combination in P. fodinatus , P. dubius , and P. churchilli . These features include highly crenulated upper molar enamel, m3 hypoconulid squared and fissured, m3 with highly crenulated talonid enamel, rounded m1–2 entoconids, centroconule on I1, and upper molars with mesostyles ( Gingerich, 1976).
Compared to the early and middle Tiffanian species of Plesiadapis , Nannodectes lynasi is much larger than Plesiadapis praecursor , P. insignis , and P. anceps . Previously, isolated plesiadapid teeth from the Goler Formation were referred to Plesiadapis cf. P. anceps ( Lofgren et al., 2008) , but molars and premolars of the three recently collected dentaries (RAM 9429, RAM 9044, and RAM 10029) indicate the size of the lower dentition of Nannodectes lynasi significantly exceeds Plesiadapis anceps ( table 6). Also, the lower premolars from these dentaries show the size of the p3 and p4 protoconid and talonid basin of p 4 in Nannodectes lynasi far surpass development of these features in Plesiadapis anceps .
The European species, P. walbeckensis , is similar to Nannodectes lynasi in size and some other features, but it has poorly developed p3–4 talonids and a fissured m3 hypoconulid with more talonid crenulations than the m3 of N. lynasi . Plesiadapis rex , is also generally similar to Nannodectes lynasi in size, but differs in having an m3 hypoconulid that is usually fissured and squared, a centroconule on I1, mesostyles on M2–3, and a mesostyle sometimes on M1 ( Gingerich, 1976). Also, Plesiadapis rex has weakly developed p3–4 talonids, compared to the relatively well-developed p3–4 talonids of Nannodectes lynasi .
Chiromyoides (particularly C. minor ) shares some features with Nannodectes lynasi as it has a rounded, broad, and unfissured m3 hypoconulid with uncrenulated talonid enamel, but its p4 lacks a talonid basin. Also, the central upper incisors of Chiromyoides are extraordinarily robust ( Gingerich, 1976), unlike Goler I1s.
In comparison to species of Nannodectes , N. lynasi is much larger than N. intermedius , N. gazini , and N. simpsoni . The lengths of the m3s and p4s in these species do not exceed 4.3 mm and 2.4 mm respectively ( Gingerich, 1976: tables A3–A 5), while in N. lynasi the lengths of m3s range from 4.32–5.43 mm and p4s from 2.52–3.05 mm ( table 6).
Nannodectes gidleyi has many similarities to N. lynasi as it has a rounded, unfissured m3 hypoconulid, lacks a centroconule on I1, and has a small talonid basin on p3–4, usually with a small but distinct cristid obliqua ( Gingerich, 1976). Also, a hypoconid and entoconid are developed on the labial and lingual edges of the posterior talonid margin of the p 4 in AMNH 17389 of N. gidleyi , a feature shared with the holotype of N. lynasi (RAM 9044). Finally, some m3s of N. gidleyi (such as AMNH 17365 and AMNH 17174) have talonid enamel crenulations that are slightly or moderately developed, indicating some overlap in this feature with N. lynasi . However, development of crenulated talonid enamel in most m3s of N. gidleyi (for example, AMNH 17171 and AMNH 17389) exceeds anything seen in N. lynasi . Also, N. gidleyi usually has mesostyles on M1 and mesostyles always on M2–3 ( Gingerich, 1976) and relatively small protoconids on p3–4. Nannodectes lynasi lacks upper molar mesostyles and has large p3–4 protoconids. Also, lower premolars and molars of N. lynasi are distinctly larger than those of N. gidleyi , although this distinction is less evident when comparing m3s ( table 6).
Nannodectes aff. N. gidleyi from Big Bend, Texas, has a p4 that is closer in size to N. lynasi , but its lower molars are smaller, about the same size as N. gidleyi ( table 6). Nannodectes aff. N. gidleyi apparently lacks a centroconule on I1 (see TMM 41365– 274 in Schiebout, 1974: fig. 16b) and has an unfissured and often rounded m3 hypoconulid lobe ( Gingerich, 1976), features shared with N. lynasi . However, Nannodectes aff. N. gidleyi has distinct upper molar mesostyles, squared hypoconulid lobes on some m3s, and highly crenulated m3 talonid enamel ( Gingerich, 1976), although the two m3s illustrated by Schiebout (1974: TMM 41365–541: fig. 16k and TMM 41366–77: fig. 17) show slightly crenulated m3 talonid enamel. In any case, N. aff . N. gidleyi and N. gidleyi are closer in morphology to N. lynasi than any other plesiadapid species.
Order PROCREODI Matthew, 1915
Family OXYCLAENIDAE Scott, 1892

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Plesiadapis
| Lofgren, Donald, Mckenna, Malcolm, Honey, James, Nydam, Randall, Wheaton, Christine, Yokote, Bryan, Henn, Lexington, Hanlon, Whitney, Manning, Stephen & Mcgee, Carter 2014 |
N. lynasi
| Lofgren & Mckenna & Honey & Nydam & Wheaton & Yokote & Henn & Hanlon & Manning & Mcgee 2014 |
N. lynasi
| Lofgren & Mckenna & Honey & Nydam & Wheaton & Yokote & Henn & Hanlon & Manning & Mcgee 2014 |
N. lynasi
| Lofgren & Mckenna & Honey & Nydam & Wheaton & Yokote & Henn & Hanlon & Manning & Mcgee 2014 |
N. lynasi
| Lofgren & Mckenna & Honey & Nydam & Wheaton & Yokote & Henn & Hanlon & Manning & Mcgee 2014 |
Nannodectes lynasi
| Lofgren & Mckenna & Honey & Nydam & Wheaton & Yokote & Henn & Hanlon & Manning & Mcgee 2014 |
N. lynasi
| Lofgren & Mckenna & Honey & Nydam & Wheaton & Yokote & Henn & Hanlon & Manning & Mcgee 2014 |
N. lynasi
| Lofgren & Mckenna & Honey & Nydam & Wheaton & Yokote & Henn & Hanlon & Manning & Mcgee 2014 |
N. lynasi
| Lofgren & Mckenna & Honey & Nydam & Wheaton & Yokote & Henn & Hanlon & Manning & Mcgee 2014 |
N. lynasi
| Lofgren & Mckenna & Honey & Nydam & Wheaton & Yokote & Henn & Hanlon & Manning & Mcgee 2014 |
Nannodectes
| Gingerich 1975 |
Nannodectes
| Gingerich 1975 |
Nannodectes
| Gingerich 1975 |
PROCREODI
| Matthew 1915 |
OXYCLAENIDAE
| Scott 1892 |