
Dipoides problematicus ( Schlosser, 1902 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.4651036 |
|
persistent identifier |
https://treatment.plazi.org/id/5862878F-E31D-FFAC-AECF-FC646C0AFC5D |
|
treatment provided by |
Felipe |
|
scientific name |
Dipoides problematicus ( Schlosser, 1902 ) |
| status |
|
Dipoides problematicus ( Schlosser, 1902) ( Fig. 2 View FIG )
NOMENCLATURAL REMARK. — Since the species problematicus is transferred from Dipoides Schlosser, 1902 to Dipoides Jaeger, 1835 (see Hugueney 1999), the correct form of the name is Dipoides problematicus ( Schlosser, 1902) .
MATERIAL EXAMINED. — Left jaw (p4-m2): PUR- 23 174. Isolated teeth: lower incisor: PUR- 23 175, CLC- 3B 84; p4: DHS- 16 214; m1-2: PUR- 23 177, DHS- 16 211; D4: PUR- 23 181; P4: CLC- 3B 83; M1-2: PUR- 23 178, DHS- 16 212; M3: DHS- 16 213. The specimen DHS- 16 216 is an undeterminable upper tooth. The worn specimen PUR- 23 176 may be a p4 or m1-2. The specimens PUR- 23 179, PUR- 23 180, DHS- 14 33 and MNA-2 51 are unspecified fragments with the typical “S”-structure of Dipoides .
MEASUREMENTS. — See Table 1. View TABLE
DESCRIPTION OF THE MATERIAL
FROM THE LOCALITY PUR-23
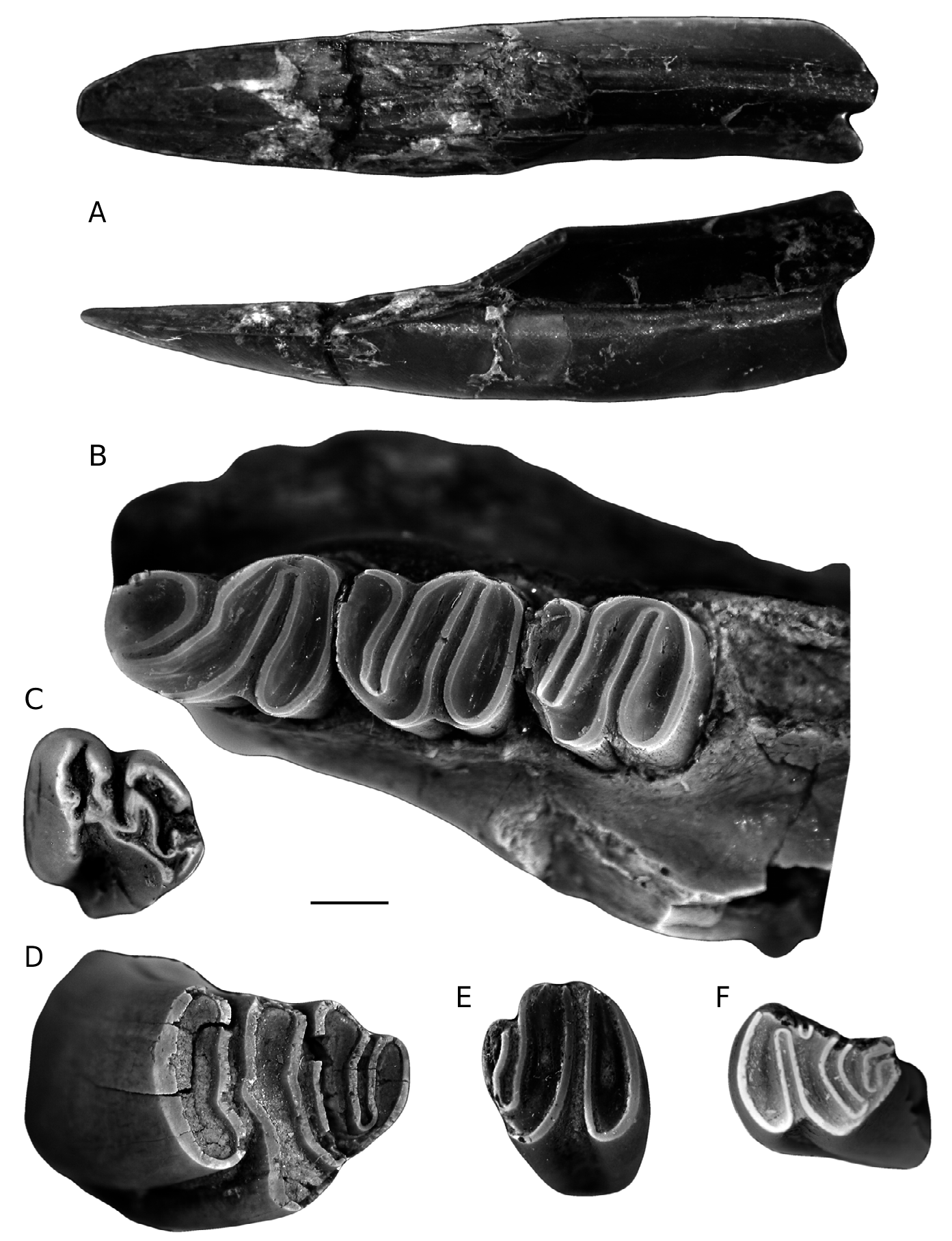
The lower incisor is convex-faced. The anterior surface is rounded and the section is oval-shaped
with a lingual central channel ( Fig. 2A View FIG ).
The p4 has the “S”-structure. It is wider posteriorly than anteriorly. The hypoflexid is longer than the mesoflexid.The mesoflexid is curved. The hypostriid is deep, in contact with the base of the crown, like the mesostriid. The paraflexid and the parastriid cannot be observed due to the wear ( Fig. 2B View FIG ).
The m1-2 has the “S”-structure. The m1 and m2 are usually described together, since they are difficult to differentiate. The m1 and m2 of PUR-23 174 are morphologically identical; the main difference is that the m2 is slightly shorter than the m1. The greatest width is posterior. The hypoflexid is longer than the mesoflexid, and they have a labial and a lingual striid respectively, which seem to connect with the base of the crown. There are no paraflexid or parastriid ( Fig. 2B View FIG ).
A single small juvenile tooth seems to correspond to an upper deciduous premolar (D4) because its crown is markedly curved forward ( Fig. 2C View FIG ).
The M1-2 has neither a metastria nor a metaflexus. The hypoflexus and the mesoflexus are relatively deep. The parastria is very much reduced. The crown is convex forward.
DESCRIPTION OF THE MATERIAL
FROM THE LOCALITY DHS-16
The p4, m1-2 and M1-2 are similar to those from PUR-23. The p4 have the paraflexid and the parastriid more developed than in the specimen from PUR-23 ( Fig. 2E View FIG ).
The M3 is lingually damaged. It is wider anteriorly than posteriorly. The hypoflexus is larger than the paraflexus, and the parastria is very short. The
hypostria is very deep. The mesoflexus is curved
and the mesostria reaches the base of the crown. Rounded metaflexus and reduced metastria. The crown is convex forward ( Fig. 2F View FIG ).
DESCRIPTION OF THE MATERIAL
FROM THE LOCALITY CLC-3B
The lower incisor is like the one from PUR-23.
The P4 is wider anteriorly than posteriorly. The crown is convex forward. The hypostria is in contact with the base of the crown. Apparently, the hypoflexus and paraflexus are in contact and form a single flexus. The parastria is shorter than the hypostria and it is attenuated at medium height of the crown. The mesostria is longer than the parastria. The metastria and metaflexus are short ( Fig. 2D View FIG ).
REMARKS
The studied teeth have the typical “S”-pattern of Dipoides , and they are difficult to determinate due to the similarities between upper and lower teeth, which sometimes only differ in the curvature of the crown.
Stehlin & Schaub (1951) and Schaub (1958) distinguish three dental patterns in the teeth of Castoridae : pentalophodont basic pattern, “S”- pattern and laminar pattern. Our specimens have the “S”-pattern, which is shared by Dipoides and Eucastor Leidy, 1858 .
Our material is medium-sized and has a simple “S”-pattern. The size and the structure agree with those of Dipoides problematicus from La Fontana, Valdecebro-3, Masada del Valle-5 and 7 ( Van de
Weerd 1976), Peralejos-E ( Adrover et al. 1988),
García-Alix A. et al.
La Gloria-5, Valdecebro-6 and Villastar ( Adrover et al. 1993; and direct comparison with material of D. problematicus from Villastar deposited in the University of Granada). The specimens from Granada Basin in general agree with, or are slightly
larger than those from the type locality of D. proble-
maticus, Salmendingen ( Schlosser 1902), but our P4 and p4 are larger than these. The structure is in general similar to that of the specimens from Salmendingen, however, some teeth have a slightly more simplified dental pattern, especially the upper
teeth. The parastria and paraflexus are very much developed (sometimes the parastria is complete) in the upper teeth from Salmendingen, much more than in our upper teeth. Our M1-2 do not show metastria nor metaflexus. The parastriid and the paraflexid are very much developed in the lower teeth from Salmendingen.
The main difference between our teeth and those of D. sigmodus (Gervais, 1859) , D. majori (Schlosser, 1903) and D. anatolicus (Özansoy, 1961) is that our specimens are smaller. Besides, D. sigmodus , D. majori and D. anatolicus have a more accentuated “S”-pattern. The “S”-pattern of D. problematicus is the most archaic. This genus is more diversified in North America than in Eurasia ( Hugueney 1999), but we did not consider American species for the determination.
Dipoides problematicus and its descendant D. sigmodus form a western and central European lineage ( Xu 1994; Hugueney 1999). Dipoides anatolicus and D. majori represent an eastern Mediterranean and Asiatic line ( Xu 1994). Following Hugueney (1999), Dipoides may either be an immigrant from America that arrived in Europe in the early Turolian or it may be derived from the Spanish Eucastor subgenus Schreuderia Aldana-Carrasco, 1992 .
DISCUSSION
Beavers are large rodents that are present in North America since the Eocene and in Eurasia since the Oligocene ( Hugueney 1999). They are very specialized mammals with concrete ecological preferences, so this group presents a great interest for paleoecological reconstructions ( Hugueney & Escuillié 1995, 1996, 1997). Semiaquatic habitats and wet preferences are attributed to this group ( Van de Weerd & Daams 1978; Van Dam & Weltje 1999). Fossorial castors, most diversified in North America, disappear at the end of the lower Miocene ( Hugueney & Escuillié 1997). Shotwell (1963) noted that Dipoides is linked to puddle zones with slow water flow. The studied localities in the Granada Basin are related with very humid conditions and fluvio-lacustrine environment (García-Alix 2006); therefore, we agree with wet and semiaquatic preferences for D. problematicus .
Until now, Castoridae were not known from
southern Spain, except for some superficial cita-
The southermost record of fossil Castoridae in Europe
tions, as indicated above. Dipoides from the Granada Basin constitutes the only record of castorids from the Turolian and Ruscinian (late Miocene and early Pliocene) in the southern Spain. Earlier records are not found in the Granada Basin because the area was submerged until the late Tortonian. In view of the very complete fossil record from the southern Spanish basins of Guadix and Crevillente ( Sesé 1989, 2006; Ruiz Bustos 1995; Martín-Suárez & Freudenthal 1998; among others), we interpret that the absence of Castoridae is real, and not due to a bias. Their presence in the Granada Basin and absence in the Guadix and Crevillente basins may be related to the different degree of humidity between these basins: the Granada Basin would have had wetter conditions in the latest Miocene-earliest Pliocene than the Guadix and Crevillente basins (Martín-Suárez et al. 2001; Minwer-Barakat 2005; García-Alix 2006). The oldest record of Castoridae in Guadix Basin, Castor sp., is from the lower Villanyian ( Sesé 1989), when the wet conditions increased (Minwer-Barakat 2005).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dipoides problematicus ( Schlosser, 1902 )
| García-Alix, Antonio, Minwer-Barakat, Raef, Martín-Suárez, Elvira & Freudenthal, Matthijs 2007 |
Schreuderia
| Aldana-Carrasco 1992 |
Eucastor
| Leidy 1858 |
Dipoides
| Jaeger 1835 |