
Ptilocerembia catherinae, Poolprasert, Pisit & Edgerly, Janice S., 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3852.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:D6781858-B9E0-4C78-A46D-971F59D56822 |
|
DOI |
https://doi.org/10.5281/zenodo.6125119 |
|
persistent identifier |
https://treatment.plazi.org/id/58376363-593F-1A41-16D1-EEE3FBCDFE23 |
|
treatment provided by |
Plazi |
|
scientific name |
Ptilocerembia catherinae |
| status |
sp. nov. |
Ptilocerembia catherinae View in CoL sp. n.
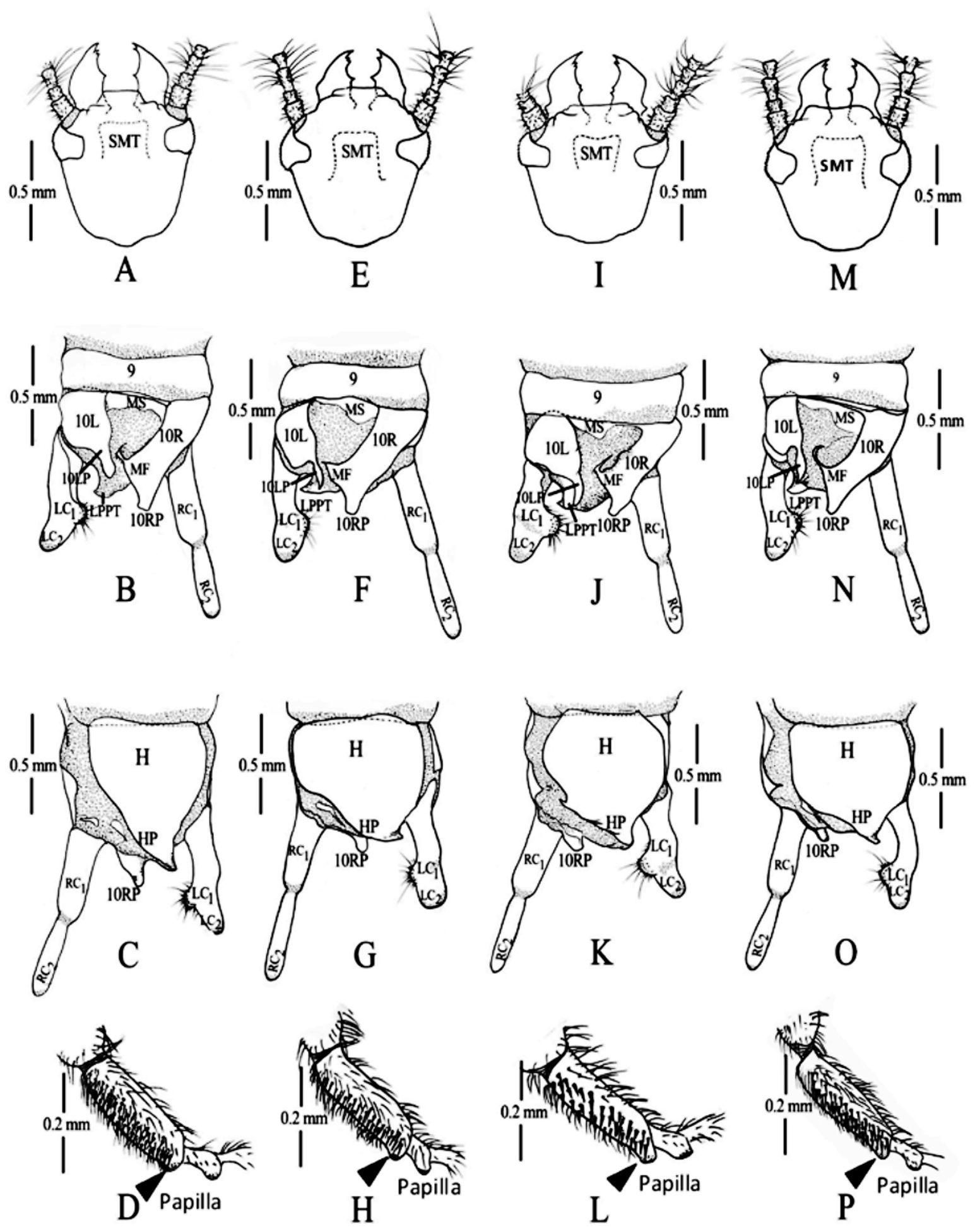
( Fig. 3 View FIGURE 3 A–3C, 4A–4B, 6C, 7I –7L)
Material examined. Holotype male ( CUMNH), Thailand, Tak province, Mae Ramat district, dry evergreen forest, 16°58.591’N, 98°31.012’E, 363 m, 20.III.2008. Seven paratype males and 15 paratype females: 1 male, 2 females ( CUMNH), same data as holotype; 2 females ( CUMNH), Mae Sod district, hill evergreen forest, 16°45.377’N, 98°31.012’E, 811 m, 20.III.2008; 1 female ( CUMNH), Mae Sod district, coniferous forest, 16°45.370’N, 98°57.056’E, 518 m 27.VI.2009; 1 female ( CUMNH), Mae Sod district, coniferous forest, 16°45.241’N, 98°56.124’E, 443 m, 13.VIII.2009; 1 male, 1 female ( CUMNH), Chiang Mai province, Sanpatong district, hill evergreen forest, 18°32.608’N, 098°31.521’E, 1237 m, 02.III.2008; 2 males, 3 females ( CUMNH), Fang district, mixed deciduous forest, 20°04.499’N 099°14.616’E, 615 m, 31.III.2008; 3 males, 4 females ( CUMNH), Mae Taeng district, hill evergreen forest, 19°18.917’N, 098°36.348’E, 1597 m, 31.III.2008; 1 female ( CUMNH), Nan province, Wiang Sa district, dry evergreen forest, 18°10.803’N, 100°58.928’E, 417 m, 22.XII.2009.
Distribution. This species is known from northern Thailand.
Etymology: The new species is named after Catherine Craig, an important and creative scientist who researches the evolution of silk and works to conserve nature by promoting local economies based on silks produced by native insects.
Description. Alate male ( Fig. 3 View FIGURE 3 A) (n = 8, mean (range) ± SD): Head width × length 2.0 (1.9–2.3) ± 0.15 × 1.8 (1.6–1.9) ± 0.11 mm; body length 16.6 (15.7–17.5) ± 0.64 mm, width 2.2 (2.0–2.4) ± 0.12 mm; forewing length 5.6 (4.8–6.1) ± 0.34 mm; hind winglength 10.5 (10.2–11.5) ± 0.32 mm.
Head capsule as broad as long ( Fig. 7 View FIGURE 7 I), blackish brown. Eyes grayish purple, paler than cranium. Preclypeal and labral membranes lavender, sclerites dark brown; mandibles dark amber, sclerotized portions of the other mouthparts dark purple, appearing concolorous to the naked eye. Molar angles of mandibles deeply pitched. Submentum trapezoidal. Antennae 30-segmented, black from base through segment 26, segments 27 to 30 white.
Thorax blackish throughout. Wings with MA forked, black with purple iridescence except for narrow, white interveinal stripes; margins of costa, radial vien and cross-veins pink. All legs entirely blackish. Hind basitarsus elongate with only one papilla, plantar setae long ( Fig. 7 View FIGURE 7 L).
Abdomen dark throughout, terminalia with 10 LP, broad, separate from 10R, 10LP broad basally, gradually arched leftward, evenly tapered to apex. 10R transverse, inner margin ending posteriorly in a blunt 10 RP, anteriorly in a dorsal sharp hook backward. Media area of inner side of 10 RP with a microspicule next to a protruding hook; EP process long, narrow, overlapping apex of an echinulated projection; LPPT somewhat long, sclerotized ( Fig. 7 View FIGURE 7 J). Right cercus with two subcylindrical segments (RC1 and RC2) ( Figs. 7 View FIGURE 7 J–7K). LC1 very short, distally echinulate, expanded and lobed; LC2 shorter, subconical, fused to LC1 ( Figs. 7 View FIGURE 7 J–7K).
Apterous female ( Fig. 3 View FIGURE 3 B) (n = 15, mean (range) ± SD): Head width × length 2.1 (2.0–2.3) ± 0.19 × 1.9 (1.8–2.0) ± 0.07 mm; body length 17.8 (16.5–18.3) ± 0.33 mm, width 2.3 (2.1–2.4) ± 0.53 mm.
Head capsule mostly blackish brown, faintly transversely clouded reddish brown between eyes. Submentum trapezoidal. Antennae 26-segmented, with basal three antennal segments golden, all other segments dark brown except distal five segments white.
Prothorax dorsally glossy dark brown, legs mostly dark brown except for entirely golden brown tibia; meso- and metathorax and their legs similar in color but tarsi are only partially medium yellowish; membranous areas between thoracic segments tan.
Abdominal terga almost all black, faintly clouded medially with golden brown; pleura creamy white forming a lateral band on each side of the abdomen; cerci entirely black; membranous areas of venter of prothorax creamy white; venter of meso- and meta-thorax and abdomen mottled dark brown; genital sternites glossy black.
Biology. Adult females were found hiding within leaf litter on the ground during the dry season in March 2008. Freshly spun silk tubes appeared pinkish rather than white, the typical color of embiopteran silk. Older silk appeared white but was usually coated with gathered materials, such as in the arboreal colony shown in Fig. 3 View FIGURE 3 C. The silk tubes in the litter, each holding one insect, fit tightly. These tubes may function as hibernacula, as if the females were aestivating until the rains began. None of the dozens of females found in leaf litter had eggs or nymphs with them. When disturbed by our collecting attempts, they would scurry rapidly into underground retreats. These tactics may provide safety against predators that dig in leaf litter. In areas near creeks or irrigated lawns, colonies were found on tree bark. These colonies appeared to be groups of nymphs, each group perhaps from one family. An egg mass uncovered in a lab colony ( Fig. 4 View FIGURE 4 ) revealed the typical style of egg clustering associated with egg-guarding and subsociality in webspinners ( Edgerly 1997). The configuration of the egg mass suggests that siblings emerge together and readily form colonies. One particularly large field colony of P. catherinae at Doi Inthanon was sequestered within a tree-hole; the silk tubes leading from this retreat to epiphytic lichens indicated that they periodically ventured forth via these tubes to graze. The behavior of this colony suggested to us that they travel back and forth from foraging sites. Further research is warranted to determine whether they cooperate to find fresh food and how they interact with colony-mates.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |