
Lernanthropus elegans, Boxshall & Bernot & Barton & Diggles & Q-Y & Atkinson-Coyle & Hutson, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4736.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:970D7D36-6D8C-4463-B9EA-D3B8E191BE72 |
|
DOI |
https://doi.org/10.5281/zenodo.3671093 |
|
persistent identifier |
https://treatment.plazi.org/id/554BDB52-7342-FFE1-5FC9-F8CB2C2FFDA8 |
|
treatment provided by |
Plazi |
|
scientific name |
Lernanthropus elegans |
| status |
sp. nov. |
Lernanthropus elegans sp. nov.
( Figs. 17–19 View FIGURE 17 View FIGURE 18 View FIGURE 19 , Fig. 20 View FIGURE 20 A–B)
Syn: Lernanthropus gisleri: Yamaguti, 1936
Type material: Holotype ♀ and allotype ♂ from Atractoscion aequidens (Cuvier, 1830) , Coffs Harbour , New South Wales; 10 December 1981; collected by K. Rohde, AM Reg. Nos. P. 103901 (Holotype ♀), P.103902 (allotype ♂) . Paratype ♀ from same host; QM Reg. No. W29488 . 2 paratype ♀♀, from A. aequidens, Coffs Harbour , New South Wales; December 1980 – January 1981; collected by K. Rohde, NHMUK Reg. No. 2018.252–253 .
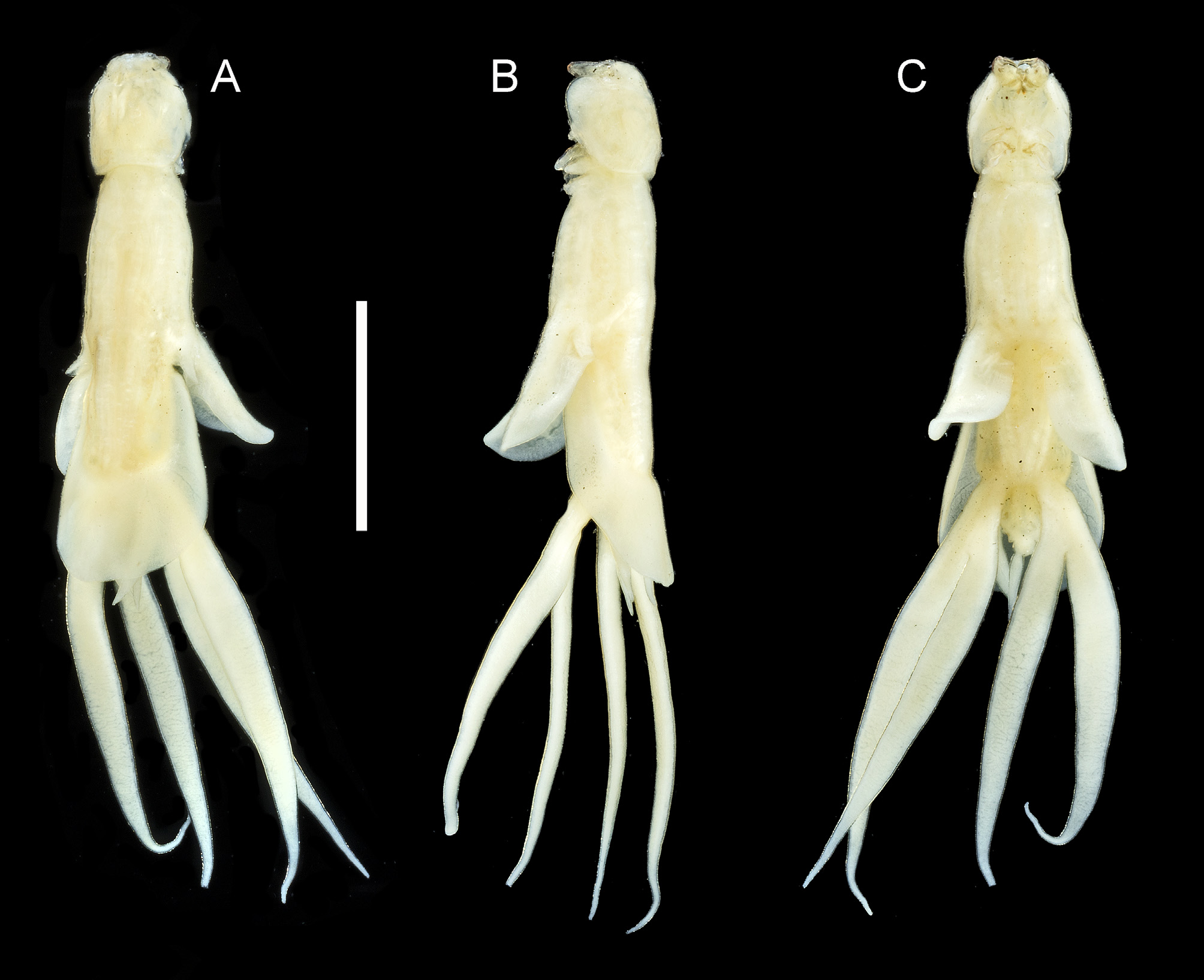
Description of female: Female body comprising cephalothorax, trunk and urosome entirely concealed beneath dorsal trunk plate ( Fig. 17 View FIGURE 17 A–C). Cephalothorax about 1.2 times longer than wide; frontal margin with weakly trilobate appearance; lateral margins more or less parallel, expanded into short, anteroventrally directed folds on either side. Trunk elongate; anterior part (second and third pedigerous somites) about 1.6 times longer than cephalothorax, becoming slightly wider posteriorly towards origin of third legs; posterior part (fourth pedigerous somite) increasing in width towards posterior margin, with dorsal trunk plate covering abdomen but with tips of caudal rami visible in dorsal view ( Figs. 17C View FIGURE 17 , 18A View FIGURE 18 ). Dorsal trunk plate with linear lateral margins increasing in width posteriorly, posterior margin evenly convex, without median indentation ( Fig. 18A View FIGURE 18 ). Urosome comprising fifth pedigerous somite, genital complex and abdomen ( Fig. 18 View FIGURE 18 A–B), all fused; dorsal surface of urosome ornamented with 6 sensillae. Each caudal ramus about 5.0 times longer than wide, tapering towards apex armed with 2 plumose setae proximally on dorsal surface, minute lateral seta located at about 87% of ramus length, and 2 small apical setae ( Fig. 18B View FIGURE 18 ). Body length of ♀ ranging from 4.64 to 4.95 mm, with a mean of 4.80 mm (based on 3 specimens).
Antennule ( Fig.18C View FIGURE 18 ) 7-segmented, segments 3 and 4 incompletely separated; setal formula: 1, 3, 2, 1, 1, 3 + ae, 7 + ae; swollen first segment and adjacent cephalothoracic surface densely ornamented with long setules. Parabasal flagellum with broad base and curved, slender distal part ( Fig. 18D View FIGURE 18 ) more than half length of antennule; swollen base lacking ornamentation. Antenna ( Fig. 18E View FIGURE 18 ) robust, comprising massive corpus, bearing papilliform element proximally on medial surface, and distal subchela unarmed but with pitted surface. Post- antennal process forming anteriorly-directed rounded lobe, densely ornamented with long setules ( Fig. 18F View FIGURE 18 ). Mandible stylet-like, armed with 8 marginal teeth distally ( Fig. 18G View FIGURE 18 ). Maxillule ( Fig. 18H View FIGURE 18 ) comprising small inner lobe tipped with 1 spiniform element and larger outer lobe with posteriorly-directed swelling at base; lobe armed with 3 unequal spiniform elements; basal swelling densely ornamented with setules. Maxilla 2-segmented: comprising proximal syncoxa (lacertus) and distal basis (brachium); basis ornamented with claw-like process distally; terminal claw ( Fig. 18I View FIGURE 18 ) armed with sharp denticles on inner surface plus 2 larger denticles proximally. Maxilliped ( Fig. 18J View FIGURE 18 ) 2-segmented; massive corpus with papilliform element on myxal surface opposing tip of subchela; distal subchela comprising fully fused endopodal segment and strongly curved terminal claw, bearing 2 small spines.
Leg 1 biramous ( Fig. 18K View FIGURE 18 ) with inflated protopod densely ornamented with long setules on anterior and lateral surfaces, armed with pinnate inner spine and slender outer seta originating close to exopod and not on a papilla: exopod 1-segmented, armed with 5 robust terminal spines (inner and outer with minute serrations on margins, middle 3 spines smooth); endopod 1-segmented, tapering distally, armed with terminal seta about as long as segment, and ornamented with spinules in distal half. Leg 2 ( Fig. 19A View FIGURE 19 ) mounted on irregularly hemispherical prominence derived from incorporated protopod; armed with outer seta on papilla: both rami 1-segmented; exopod armed with 4 distal margin spines; endopod armed with small apical seta less than half length of segment; both rami ornamented with patches of spinules. Leg 3 located ventrolaterally at rear of third pedigerous somite, endopod forming long fleshy, shoehorn-shaped, lamella, directed laterally: third legs separate along midline; slightly outer protrusion of margin at base of lamella possibly representing reduced exopod; leg 3 armed with dorsal outer basal seta ( Fig. 17 View FIGURE 17 A–C). Leg 4 bilobate; inner and outer lobes elongate, protruding well beyond free posterior margin of dorsal trunk plate ( Fig. 17 View FIGURE 17 A–C); outer protopodal seta located dorsally at base of limb. Leg 5 ( Fig. 18 View FIGURE 18 A–B) forming lanceolate process extending well beyond tips of caudal rami and beyond rear margin of dorsal trunk plate; outer protopodal seta present dorsally near base of process; ornamented with surface sensillae proximally.
Description of male: Body smaller than female ( Fig. 19B View FIGURE 19 ), total length about 2.43 mm. Cephalothorax comprising about 34% of total body length, with convex lateral margins becoming wider posteriorly; dorsal cephalic shield about 1.4 times longer than maximum width. Frontal area of cephalothorax carrying antennule and antennae, defined by weak indentation. Trunk slender, comprising all fused post-cephalothoracic somites ( Fig. 19B View FIGURE 19 ), including urosome. Ventral surface of genital area ornamented with setules ( Fig. 19C View FIGURE 19 ). Anal somite unornamented; bearing paired caudal rami; each ramus elongate, about 5.0 times longer than maximum width, armed with 2 plumose setae proximally on dorsal surface, 1 short lateral seta located at 84% of ramus length, plus 2 apical setae.
Antennule 7-segmented as in female. Parabasal flagellum as in female. Antenna ( Fig. 19D View FIGURE 19 ) comprising long, slender corpus and distal subchela terminating in strongly recurved claw: corpus armed with papilliform element proximally on medial surface; subchela with small process in area of proximal articulation plus larger claw like process near middle. Postantennal process and mandible as in female. Oral cone ornamented with scattered spinules ( Fig. 19E View FIGURE 19 ). Maxillule bilobate; smaller lobe with blunt apical element, larger lobe armed with 3 unequal distal elements and ornamented with spinules ( Fig. 19E View FIGURE 19 ). Maxilla as in female. Maxilliped ( Fig. 19F View FIGURE 19 ) comprising robust corpus with small myxal process and ornamented with patches of spinules proximally on medial surface, and distal subchela armed with 2 spines.
Leg 1 robust ( Fig. 20A View FIGURE 20 ), basis armed with slender outer seta and stout inner spine; exopod 1-segmented, broader distally, ornamented with spinules as figured and armed with 5 distal spines, all spinulate; endopod 1-segmented, tapering distally, distally and armed with spinulose apical seta just shorter than segment; segment ornamented with long spinules distally. Leg 2 ( Fig. 20B View FIGURE 20 ) lacking intercoxal sclerite; basis with slender outer seta; both rami 1-segmented; exopod swollen distally, armed with 3 spines plus a hemispherical swelling and other spinous structures distally, plus rows of spinules on distal surface; endopod tapering distally and armed with long unilaterally spinulose seta apically; seta about as long as segment; surface of segment ornamented with spinules along medial surface. Leg 3 ( Fig. 19G View FIGURE 19 ) biramous, armed with basal seta dorsally at base of limb; exopodal lobe long and cylindrical, directed posterolaterally from ventrolateral origin on trunk; endopodal lobe about one third length of exopodal lobe. Leg 4 ( Fig. 19B View FIGURE 19 ) biramous with both rami forming elongate lobes, outer basal seta present dorsally at base of limb. Leg 5 represented by single pinnate seta located laterally on urosome ( Fig. 19C View FIGURE 19 ).
Remarks: The new species was found on Atractoscion aequidens , a member of the family Sciaenidae , and this family serves as host to about 20 species of Lernanthropus in total ( Table 3). Most of the species listed in Table 3 are exclusively parasitic on sciaenid hosts, but a few, such as L. gisleri and L. leidyi have occasionally been reported from non-sciaenids. In addition, L. pomatomi , L. pupa and L. paenulatus were all originally described from nonsciaenid hosts but each has been reported on a single occasion from a sciaenid host. Detailed comparison of the new species with all of these other species found on sciaenid hosts reveals some significant differences as well as shared character states.
The new species, L. elegans sp. nov., can be readily distinguished from L. huamani Luque & Farfán, 1990 , L. longipes Wilson, 1932 , L. pacificus Oliva & Durán, 1982 , and L. paralonchuri Luque, Bruno & Covarrubias, 1989 , by the length of the lobes of leg 4, which are shorter than the body in the new species but markedly longer than the entire body in these four species. Another very distinctive species is L. grassei . This is an elongate, slender species which has very unequal lobes on leg 4: the exopodal lobe is almost twice the length of the endopodal lobe. This contrasts with the more-or-less equal lobes of the new species. The South American species L. cynoscicola has an unusually short dorsal trunk plate so that the tip of the abdomen and the entire length of the caudal rami are visible in dorsal view, whereas in L. elegans sp. nov. the dorsal trunk plate is more extensive, concealing the entire urosome and all but the tips of the caudal rami.
Lernanthropus barnardi Capart, 1959 and L. nunesi Capart, 1959 are both known only from their inadequate original descriptions in Capart (1959). However, the former can be distinguished by the relatively short lanceolate lobes of leg 4 which extend beyond the posterior margin of the dorsal trunk plate only by about 30% of their total length, whereas in the new species about 70% of the length of these lobes is visible in dorsal view. The latter species, L. nunesi , has elongate fourth leg lobes but differs from the new species in having a very reduced leg 5 compared to the large lobate leg 5 of the new species. The original description of L. leidyi was also inadequate, but both sexes of this species were redescribed by Luque & Paraguassú (2003). Comparing the new species with this redescription of L. leidyi allows us to distinguish between them by the length of dorsal trunk plate, which is longer than the anterior part of the trunk in the new species but distinctly shorter in L. leidyi , and by the fifth leg which is elongate in the new species but short and subcylindrical in L. leidyi , according to Luque & Paraguassú (2003). The fifth legs of L. pagodus Krøyer, 1863 appear to be reduced or absent, since Krøyer (1863: Tab VIII, Fig. 2c View FIGURE 2 ) does not figure them in his ventral habitus drawing. This species also differs in having a broad dorsal trunk plate (about 2.0 times wider than the cephalothorax) with a median indentation in its posterior margin, whereas L. elegans sp. nov. has a more slender dorsal trunk plate (only about 1.5 times wider than the cephalothorax) with an evenly convex posterior margin.
The new species shares a distinctive set of character states with the remaining eight species from Table 2 View TABLE 2 , including the widely distributed L. gisleri . This character set includes: 7-segmented antennule with the segment bearing the subapical aesthetasc separate from the apical segment (these segments are fused in many Lernanthropus species); the proximal segment of the antennule is fused to a swelling on the surface of the cephalothorax and both the segment and swelling are densely ornamented with long setules; the maxilla bears a curved process on the basis just proximal to the base of the apical claw; the protopod of leg 1 is densely ornamented with long setules; leg 3 forms a large, (postero-) laterally directed lamella; the rami of leg 4 are both elongate; leg 5 forms an elongate process; and the caudal rami are elongate. Detailed comparisons are required to separate L. elegans sp. nov. from these eight species: L. gisleri , L. capistroides Olivier & van Niekerk, 1995 , L. cruciatus Pillai, 1962 , L. francai Nuñes-Ruivo, 1962 , L. indefinitus Koyuncu, Castro-Romero & Karaytug, 2012 , L. otolithi Pillai, 1963 , L. pami Tripathi, 1962 , and L. sciaenae Gnanamuthu, 1947 .
Lernanthropus pami is included in this group but, unfortunately, only limited comparisons are possible because this species is very poorly known and its original description contains major mistakes (cf. Pillai, 1985). The information available from Tripathi (1962) indicates that the fifth legs (erroneously interpreted as a third lobe of a “trifoliate” leg 4) are elongate but extend only to about the mid-level of the caudal rami. In contrast, L. elegans sp. nov. has lanceolate fifth legs that extend well beyond the distal tips of the caudal rami. Additionally, the tips of the caudal rami only just reach the posterior margin of the dorsal trunk plate in L. pami but extend well beyond this margin in the new species. Tripathi (1962) remarked that L. pami was similar to L. sciaenae described by Gnanamuthu (1947), probably because Gnanamuthu had also misinterpreted leg 4 as trifoliate. His figure of the antennule of L. sciaenae shows the setular ornamentation on the proximal segment, even though the description is lacking in other details. Lernanthropus sciaenae resembles L. pami in the proportional lengths of the fifth legs and caudal rami, and in the extent of the caudal rami relative to the margin of the dorsal trunk plate, and differs from L. elegans sp. nov. in these features. We propose to treat L. pami as a species inquirendum, but it is also possible that L. pami is a junior synonym of L. sciaenae .
The original description of L. capistroides was supported only by scanning electron micrographs ( Olivier & van Niekerk, 1995). These images provide considerable detail of the appendages: for example, the maxilla of the female carries a curved claw (canna in Olivier & van Niekerk, 1995) on the inner margin of the basis plus a thinwalled process (clavus in Olivier & van Niekerk, 1995) on the base of the terminal claw (calamus in Olivier & van Niekerk, 1995). This armature is shared with L. elegans sp. nov. However, Olivier & van Niekerk (1995) did not provide an illustration of the habitus, which renders comparisons with older descriptions problematic. The most readily observable difference between L. capistroides and the new species is the caudal rami, which do not reach the posterior margin of the dorsal trunk plate in the former but extend well beyond this margin in the latter. The mean body length of L. capistroides is given as 7.2 mm by Olivier & van Niekerk (1995), of which the dorsal cephalic shield comprised 1.3 mm and the dorsal trunk plate 3.2 mm. We can calculate that the mean length of the anterior trunk was therefore 2.7 mm. This is about 2.1 times longer than the cephalothorax which differs from the new species, in which the anterior trunk is only 1.6 times longer than the cephalothorax.
Lernanthropus francai was first described from off the coast of Angola ( Nuñes-Ruivo, 1962) and has only been reported once since, in a list of copepods found on Vietnamese fishes ( Kazachenko et al., 2014). It is known only from the female which exhibits many detailed similarities to L. elegans sp. nov. For example, both species have a densely setulose outer expansion of the protopodal part of leg 1 and the endopod of this leg bears a single apical seta that is about as long as the ramus. Both also have the setulose, posteriorly-directed proximal lobe on the maxillule. The key difference between these species is the extent of the caudal rami, the tips of which do not reach the posterior margin of the dorsal trunk plate in L. francai , but extend beyond it in L. elegans sp. nov. They can also be distinguished by the relative lengths of the lanceolate rami of leg 4 relative to the dorsal trunk plate: in L. francai these rami extend beyond the posterior margin of the plate by slightly more than half of their entire length whereas in the new species about 75% of their length is visible in dorsal view, extending beyond the margin.
The Indian species L. cruciatus has a body length of over 16 mm ( Pillai, 1985) and is instantly recognizable by its enormous third legs, which are laterally directed and about equal in length to the cephalothorax and anterior trunk combined. The dorsal trunk plate of L. cruciatus is also very well developed, comprising just over half (53%) of the entire body length. In contrast, L. elegans sp. nov. has third legs that are slightly shorter than the anterior trunk region alone, and its dorsal trunk plate comprises only 42% of body length.
The slender body shape of the female of L. elegans sp. nov. contrasts with the rather robust body form of L. otolithi . The latter has an almost circular dorsal trunk plate which is slightly wider than long whereas in the former the plate is about 1.3 times longer than wide and has tapering linear lateral margins that expand posteriorly. The ventrally-produced lateral margins of the dorsal cephalic shield of L. otolithi project anteriorly generating a trilobate frontal margin whereas the frontal margin is straight with rounded corners in L. elegans sp. nov.
The original description of L. indefinitus ( Koyuncu et al., 2012) combines some excellent illustrations of particular limbs (such as the antennule and legs 1 and 2) with inadequate illustrations which fail to show important features (such as the caudal rami, from which most of the caudal setae are missing). In female L. indefinitus the fifth leg lobes do not reach the tips of the caudal rami whereas in L. elegans sp. nov. they extend well beyond the tips of the rami. The caudal rami themselves are about 3 times longer than wide in L. indefinitus compared to about 5 times longer than wide in the new species.
Finally, the new species can be differentiated from L. gisleri by the length of the caudal rami and fifth legs relative to the free posterior margin of the dorsal trunk plate: in L. gisleri the caudal rami are completely concealed in dorsal view, hidden beneath the dorsal trunk plate while the fifth legs reach to the posterior margin of the dorsal trunk plate and the tips may be visible dorsally in some specimens. In contrast, in L. elegans sp. nov. the tips of the caudal rami and the distal 25% of the fifth legs extend beyond the margin of the plate and are visible in dorsal view. The structure and armature of the limbs are very similar in these two species. The antennules are 7-segmented in both and the setation is almost identical. The absence of setular ornamentation on segment 1 in Kabata’s figure (1979b: fig. 1058) is significant since it is very conspicuous in L. elegans sp. nov. and would not be easy to overlook. Both species possess a hirsute posterior swelling at the base of the maxillule. Legs 1 and 2 are also very similar in the two species. The only difference in leg 1 is the extent of the setular ornamentation, which is much more extensive in L. elegans sp. nov. In leg 2 both rami are ornamented with surface spinules in the new species but, according to Kabata (1979b), only the endopod carries any spinules in L. gisleri .
The four Lernanthropus females from Sciaena sp. identified by Yamaguti (1936) as L. gisleri are misidentified, as already noted by Koyuncu et al. (2012). Although not described in full by Yamaguti (1936), this material appears to belong to the new species, L. elegans sp. nov., and it seems possible that the material of L. gisleri listed, but not illustrated by Song and Chen (1976) from Johnius dussumieri (Cuvier, 1830) (as Sciaena dussumieri ) caught in China, might also be attributed to the new species.
Although not parasitic on a sciaenid host, L. antofagastensis Castro-Romero & Baeza-Kuroki, 1985 is included in our comparisons. This species is parasitic on a haemulid, Anisotremus scapularis (Tschudi, 1846) , found in Chilean coastal waters and it shares many character states with L. elegans sp. nov. including: the swollen proximal segment of the antennule with a dense ornamentation of setules, the swollen and densely setulose base of the parabasal flagellum, the armature on the basis of the maxilla, the long cylindrical fifth legs and the elongate caudal rami. However, these two species can be distinguished by the relative lengths of the fifth legs which extend well beyond the tips of the caudal rami in L. elegans sp. nov., but only to the base of the caudal rami in L. antofagastensis .




| AM |
Australian Museum |
| QM |
Queensland Museum |
| NHMUK |
Natural History Museum, London |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |