
Sagum lativentris ( Heller, 1865 ), 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4736.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:970D7D36-6D8C-4463-B9EA-D3B8E191BE72 |
|
DOI |
https://doi.org/10.5281/zenodo.3671083 |
|
persistent identifier |
https://treatment.plazi.org/id/554BDB52-7333-FF98-5FC9-F9352859F890 |
|
treatment provided by |
Plazi (2020-02-17 07:18:21, last updated 2024-11-26 03:30:40) |
|
scientific name |
Sagum lativentris ( Heller, 1865 ) |
| status |
comb. nov. |
Sagum lativentris ( Heller, 1865) n. comb.
( Figs. 44–46 View FIGURE 44 View FIGURE 45 View FIGURE 46 )
Syn: Lernanthropus lativentris Heller, 1865
Sagum lativentris: Pattipeiluhu & Gill, 1998
non Lernanthropus lativentris: Pillai, 1985
Material examined: 3♀♀ from Lutjanus johnii (Bloch, 1792) , Darwin (Outer Harbour), 07 October 2012, collected by B.K. Diggles; MAGNT Reg. No. Cr019249. 1♀ from L. johnii , Cape Hotham, Northern Territory; 03 January 2014; collected by D.P. Barton; QM Reg. No. W29504. 1♂ from L. johnii, Lorna Shoal, Timor Sea , Northern Territory; 15 August 2013; collected by D.P. Barton; MAGNT Reg. No. Cr019250. 1♂ from L. johnii, Melville Island, Northern Territory; 12 September 2013; collected by D.P. Barton; QM Reg. No. W29505. 1♀ from L. johnii , Cape Hotham, Northern Territory; 03 January 2014; collected by D.P. Barton. 1♂ from L. johnii, Melville Island, Northern Territory; 12 September 2013; collected by D.P. Barton; NHMUK Reg. No. 2018.303.
1♀ from Lutjanus russellii (Bleeker, 1849) , Darwin (Outer Harbour), 07 October 2012, collected by B.K. Diggles; NHMUK Reg. No. 2018.304 .
4♀♀ from Lutjanus carponotatus (Richardson, 1842) , Bynoe Harbour , Northern Territory, 28 August 2012, collected by B.K. Diggles; NHMUK Reg. No. 2018.305 .
Comparative material examined: 3♀♀, 2♂ from Lutjanus vitta (Quoy & Gaimard, 1824) (as Mesoprion phaiotaeniatus ), Ambon, Indonesia; summer 1993, collected by S. Pattipeiluhu; NHMUK Reg. No. 1994.711–715 .
Differential diagnosis: Cephalothorax about 1.1 times longer than wide with almost linear lateral margins narrowing anteriorly towards short frontal margin ( Fig. 44 View FIGURE 44 A–C); lateral margins of dorsal cephalothoracic shield projecting ventrally, with evenly convex free margin in lateral view ( Fig. 44B View FIGURE 44 ). Anterior part of trunk (second and third pedigerous somites) about 2.1 times wider than long with distinct anterolateral shoulders; becoming slightly wider posteriorly and with short tapering posterolateral processes; posterior part (fourth pedigerous somite) covered by dorsal trunk plate. Dorsal trunk plate subcircular, with convex lateral and strongly convex posterior margins ( Fig. 44 View FIGURE 44 A–C); entirely concealing urosome and fourth legs in dorsal view. Urosome comprising fifth pedigerous somite, genital complex and abdomen, all fused ( Fig. 45A View FIGURE 45 ). Genital complex slender, with conspicuous paired gonopores dorsolaterally and with paired copulatory pores located posterolaterally on ventral surface (arrowed in Fig. 45A View FIGURE 45 ); dorsal surface of genital complex ornamented with 2 pairs of sensillae located between gonopores. Egg sacs loosely coiled beneath dorsal trunk plate and partly concealed laterally by third legs. Paired caudal rami elongate, about 2.8 times longer than maximum width; broadest in mid-section. Caudal rami each with 2 plumose dorsal setae; distal dorsal seta located at 44% of length of ramus, small outer seta located at about 60% of ramus length, plus 2 apical setae ( Fig. 45A View FIGURE 45 ): tips of caudal rami reaching beyond mid-length of lobes of leg 4 ( Fig. 45B View FIGURE 45 ).
Antennule unsegmented, armed with 7 setae proximally and 12 setae plus 2 aesthetascs around apex ( Fig. 45C View FIGURE 45 ). Parabasal flagellum absent. Antenna with robust proximal segment armed with conical process on medial margin; distal subchela armed with 2 rounded processes plus smaller process in articulation with proximal segment ( Fig. 45D View FIGURE 45 ). Postantennal process rounded ( Fig. 45E View FIGURE 45 ). Mandible ( Fig. 45F View FIGURE 45 ) stylet-like with 8 marginal teeth at apex. Maxillule bilobed, smaller lobe tipped with 1 spiniform element; larger lobe tipped with 3 unequal spiniform elements and ornamented with patch of hair-like setules ( Fig. 45G View FIGURE 45 ). Maxilla ( Fig. 45H View FIGURE 45 ) with short tapering syncoxa; basis with apical claw ornamented with row of denticles, plus blunt subapical process. Maxilliped corpus with irregular myxal surface but no obvious processes; subchela armed with inner seta on concave margin ( Fig. 46A View FIGURE 46 ).
Leg 1 with coxa fused to somite and incompletely separated from basis; members of leg pair joined by intercoxal sclerite ( Fig. 46B View FIGURE 46 ). Each leg biramous with outer plumose seta and short, hirsute inner spine on basis; exopod 1-segmented, armed with 5 robust terminal spines with denticulate margins; endopod 1-segmented, smoothly tapering to unarmed apex; surface unornamented. Leg 2 ( Fig. 46C View FIGURE 46 ) carried on inflated subspherical prominence derived from incorporated protopod armed with outer plumose seta: biramous, with both rami 1-segmented and both with wrinkled cuticular surface; exopod armed with 5 small distal spines; endopod incompletely separated basally, unarmed. Leg 3 located on ventral surface of third pedigerous somite, forming bilobed fleshy lamella, large outer lobe held vertically and directed posteriorly, reaching about to middle of dorsal trunk plate; smaller inner lobe about one third length of outer lobe; held vertically and extending posteromedially towards midline but legs separate along midline. Leg 4 bilobate ( Fig. 45B View FIGURE 45 ); inner and outer lobes elongate, flattened and tapering slightly from wider base; distal part irregularly bent and usually concealed beneath dorsal trunk plate, but when fully extended and straight, reaching beyond free posterior margin of trunk dorsal plate. Leg 5 represented by minute papilla carrying short apical seta (arrowheads in Fig. 45A View FIGURE 45 ). Body length of ♀ ranging from 2.18 to 3.20 mm, with a mean of 2.72 mm (based on 9 specimens).
Description of male. Body smaller than female, comprising cephalothorax covered with dorsal shield and subrectangular trunk, separated by narrow neck region ( Fig. 46D View FIGURE 46 ); dorsal surface of shield and trunk both densely ornamented with minute papillae. Cephalothorax about 1.4 times longer than wide with angular posterolateral corners and convex posterior margin. Trunk about 1.3 times longer than wide; lateral margins sinuous; posterior margin with indentations at base of leg 4 and medial to paired genital apertures: anal somite fused to trunk, wider than long, bearing paired caudal rami plus medial tapering process ( Fig. 46E View FIGURE 46 ). Caudal rami irregularly cylindrical; bearing 2 dorsal setae in proximal half, lateral seta located at two-thirds of ramus length, plus 2 unequal apical setae.
Antennule unsegmented and armed as in female. Parabasal flagellum curved, with broad base tapering towards slender tip ( Fig. 46F View FIGURE 46 ). Antenna, mandible and maxillule as in female. Maxilla similar to female but subapical process on basis smaller ( Fig. 46G View FIGURE 46 ). Maxilliped with unisensillate papilla on myxal surface; subchela with small process distal to tiny seta on concave margin; apical claw ornamented with row of tiny spinules at base and with surface striations and pits towards apex ( Fig. 46H View FIGURE 46 ).
Leg 1 as in female except vestigial setal element on tip of endopod better developed ( Fig. 46I View FIGURE 46 ). Leg 2 ( Fig. 46J View FIGURE 46 ) with better defined exopod with complex surface ornamentation; endopod lobate, unarmed. Leg 3 biramous ( Fig. 46D View FIGURE 46 ), with each ramus forming elongate cylindrical lobe; armed with outer protopodal seta situated on small papilla located dorsally at base. Leg 4 ( Fig. 46D View FIGURE 46 ) similar to leg 3, also armed with outer protopodal seta situated on small papilla located dorsally at base. Leg 5 absent. Leg 6 represented by paired genital apertures, unarmed. Body lengths of measured ♂♂ 1.59 and 1.66 mm.
Distribution: The original description of Sagum lativentris (as Lernanthropus lativentris ) was based on females found on Lutjanus vitta (Quoy & Gaimard, 1824) (as Mesoprion phaiotaeniatus ) caught off Java, Indonesia ( Heller, 1865). Pattipeiluhu & Gill (1998) reported this species from the same host caught off Ambon, Indonesia, but other records of this species from India have been shown to refer to a different species (see Remarks below). The report of S. lativentris (as L. lativentris ) on the gempylid Thyrsitoides marlayi Fowler, 1929 from Mozambique ( Reimer, 1986) is so unusual as to raise considerable doubt over the identification. [The only Sagum species reported from a gempylid host is S. foliaceum known from New Zealand waters ( Hewitt, 1968).] The present account is the first report of S. lativentris n. comb. from Australian waters.
Remarks: Pillai (1985) reported a species under the name of “ Lernanthropus lativentris ” from Lethrinus harak (Forsskål, 1775) (as L. rhodopterus ) and Lutjanus vitta in Indian waters. Kabata (2005) concluded that the material identified and described by Pillai (1985) as L. lativentris in his monograph was misidentified, and he established L. pillaii Kabata, 2005 as a new species to accommodate Pillai’s material. We follow Kabata in recognizing that Heller’s L. lativentris is valid and distinct, but this species remained poorly known. It is unfortunate that Heller’s type material was not ovigerous, since the morphology of the egg sacs indicates that this species belongs in Sagum . The discovery of ovigerous material from the type host Lutjanus vitta (as Mesoprion phaiotaeniatus ) collected in Ambon, Indonesia revealed the nature of the loosely coiled egg sacs, so this discovery was reported by Pattipeiluhu & Gill (1998) under the combination Sagum lativentris , although the new combination status was not made explicit.
There are another two very similar species currently placed in Lernanthropus , L. caesionis Yamaguti, 1954 and L. sanguineus Song, in Song & Chen, 1976 , which also appear to belong in Sagum . The former species was described by Yamaguti (1954) based on two females taken from the gills of Caesio cuning (Bloch, 1791) (as C. kuning ) caught off Sulawesi, Indonesia. A complete description of this species is not available but in addition to the habitus of the female, the unarmed endopod of leg 1 and the leaf-like shape of the caudal rami are typical of a cluster of Sagum species (see Table 4 View TABLE 4 ). We here transfer L. caesionis to Sagum as S. caesionis ( Yamaguti, 1954) n. comb. We note that S. caesionis is very similar to S. gurukun Uyeno & Naruse, 2018 , but can be distinguished by the length of the rami of leg 4. Finally, L. sanguineus was based on study of two females collected from Lutjanus sanguineus (Cuvier, 1828) caught at Sanya, Hainan Dao, China ( Song & Chen, 1976). The females were not ovigerous but the only subsequent record of this copepod was under the name Sagum sanguineus ( Song, 1976) and came from Vietnamese waters. This report was based on females from Lutjanus johnii ( Kazachenko et al., 2014) and the name used was not explicitly identified as constituting a new combination, but the transfer to Sagum is confirmed below.
After these generic transfers, Sagum now comprises 18 species ( Table 4 View TABLE 4 ). These three newly transferred species were not included in the comparisons made by Uyeno & Naruse (2018) when they established two new species of Sagum from the Ryukyu Islands, Japan, nor were they considered in the brief discussion in Kazachenko et al. (2017) who established S. vietnamensis Kazachenko, Kovalev, Nguyen & Ngo, 2017 . The key to species of the genus provided by Uyeno & Naruse (2018) needs to be updated, but here we provide a Table comparing key character states of all species.
The material of S. lativentris collected from Lutjanus johnii in Australian waters has smooth linear lateral margins of the cephalothorax and tapering fourth legs, the tips of which may just extend beyond the posterior margin of the dorsal trunk plate when straightened. There are nine species that share these two character states ( Table 4 View TABLE 4 ). These same species all have a leaf-like caudal ramus, with the exception of S. vietnamensis which has cylindrical caudal rami that are about 4.4 times longer than wide according to Kazachenko et al. (2017). Sagum lativentris has caudal rami that are about 2.8 times longer than wide and can be readily separated from S. caesionis and S. gurukun , which have short caudal rami (only 1.4 to 1.5 times longer than wide). The setation on the caudal ramus also provides useful characters: in S. bitaro Uyeno & Naruse, 2018 , S. folium , and S. paracaesionis Izawa, 2014 , the 2 dorsal caudal setae are located in the distal half of the ramus, whereas in S. lativentris these 2 setae originate in the proximal half. Sagum lativentris differs from S. foliaceum in body proportions: so the cephalothorax and anterior trunk combined are longer than the dorsal trunk plate in S. lativentris but markedly shorter in S. foliaceum . In addition the lobes of leg 4 extend well beyond the posterior margin of the dorsal trunk plate in the latter, but only the tips are visible in dorsal view in the former. Finally, S. lativentris and S. sanguineus are very similar morphologically and both use lutjanids as hosts. Indeed, both parasites have been recorded from the same host, L. johnii (present account; Kazachenko et al. 2014). The best characters to differentiate between them relate to the fourth legs and caudal rami. In S. lativentris the fourth legs have somewhat flaccid tips and are often bent, but when straightened the tips are visible beyond the posterior margin of the dorsal trunk plate, whereas in S. sanguineus the fourth leg lobes are more linear and the distal third of each is visible in dorsal view. The caudal rami in both species are about 2.7 to 2.8 times longer than wide but in S. lativentris the distal of the two dorsal setae is located at about 44% of the ramus length while in S. sanguineus it is located at about 30% of the ramus length. In addition, the tips of the caudal rami reach beyond the mid-length of the fourth leg lobes in S. lativentris ( Fig. 45B View FIGURE 45 ) but do not reach the mid-length in S. sanguineus ( Fig. 47C View FIGURE 47 ).
Heller, C. (1865) Crustaceen. In: Reise der Osterreichischen Fregatte Novara um die Erde in den Jahren 1857, 1858, 1859, Zoologie, 2, 1 - 280, pls. 1 - 25. https: // doi. org / 10.5962 / bhl. title. 1597
Hewitt, G. C. (1968) Some New Zealand parasitic Copepoda of the family Anthosomidae. Zoology Publications from Victoria University of Wellington, 47, 1 - 31.
Izawa, K. (2014) Some new and known species of the Lernanthropidae (Copepoda, Siphonostomatoida) parasitic on the branchial lamellae of Japanese actinopterygian fishes, with revision of two known species of the family and discussion on the insemination mode in the Siphonostomatoida. Crustaceana, 87, 1521 - 1558. https: // doi. org / 10.1163 / 15685403 - 00003373
Kabata, Z. (2005) Lernanthropus lativentris Heller, 1865 (Copepoda, Lernanthropidae) of Pillai (1985) recognised as a new species. Acta Parasitologica, 50, 352 - 354.
Kazachenko, V. N., Kovaleva, N. N., Nguyen, V. T. & Ngo, H. D. (2014) Taxonomic review of the parasitic copepod (Crustacea: Copepoda) fish in Vietnam. Scientific Journal of Dalrybvtuz, 31, 20 - 30.
Kazachenko, V. N., Kovaleva, N. N., Nguyen, V. T. & Ngo, H. D. (2017) Three new species and one new genus of parasitic copepods (Crustacea: Copepoda) from fishes of the South China Sea. Russian Journal of Marine Biology, 43, 264 - 269. https: // doi. org / 10.1134 / S 1063074017040058
Pattipeiluhu, S. M. & Gill, M. E. (1998) Ectoparasites of Coral Reef Fishes and Their Value as Biological Indicators of Pollution. Cakalele, 9, 25 - 29.
Pillai, N. K. (1985) Fauna of India. Parasitic copepods of marine fishes. Zoological Survey of India, Calcutta, 900 pp.
Reimer, L. W. (1986) Parasitic copepods of fishes from the coast of Mozambique. Wiadomosci Parazytologiczne, 32, 505 - 506.
Song, D. & Chen, G. (1976) Some parasitic copepods from marine fishes of China. Acta Zoologica Sinica, 22, 406 - 424.
Uyeno, D. & Naruse, T. (2018) Two new species of Sagum Wilson, 1913 (Copepoda: Siphonostomatoida: Lernanthropidae) parasitic on reef fishes off the Ryukyu Islands. Systematic Parasitology, 95, 893 - 904. https: // doi. org / 10.1007 / s 11230 - 018 - 9810 - 4
Yamaguti, S. (1954) Parasitic copepods from fishes of Celebes and Borneo. Publications of the Seto Marine Biological Laboratory, 3, 375 - 398. https: // doi. org / 10.5134 / 174483
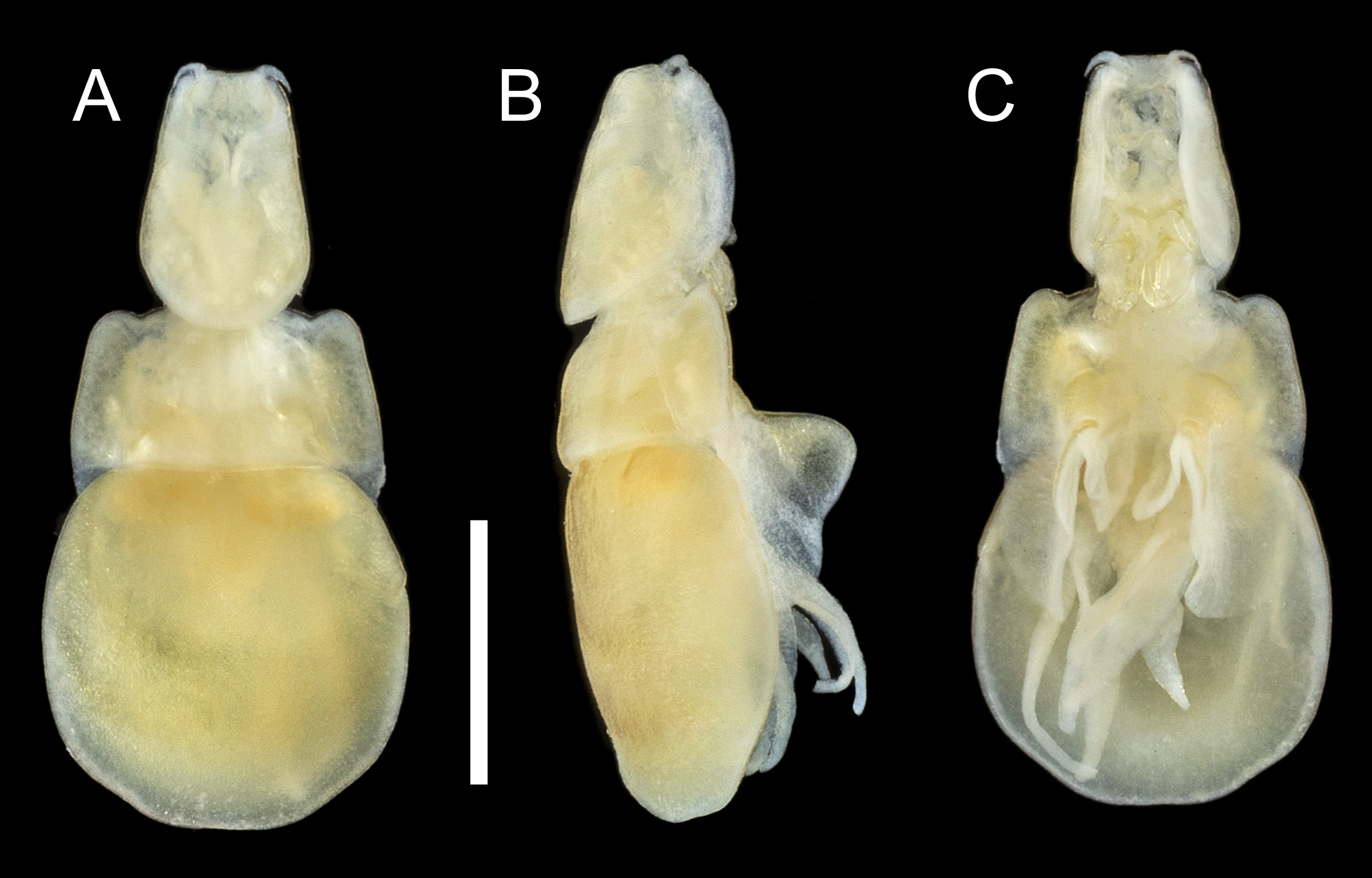
FIGURE 44. Sagum lativentris (Heller, 1865) n. comb., adult ♀. A, habitus, dorsal; B, habitus, lateral; C, habitus, ventral. Scale bar 1 mm.
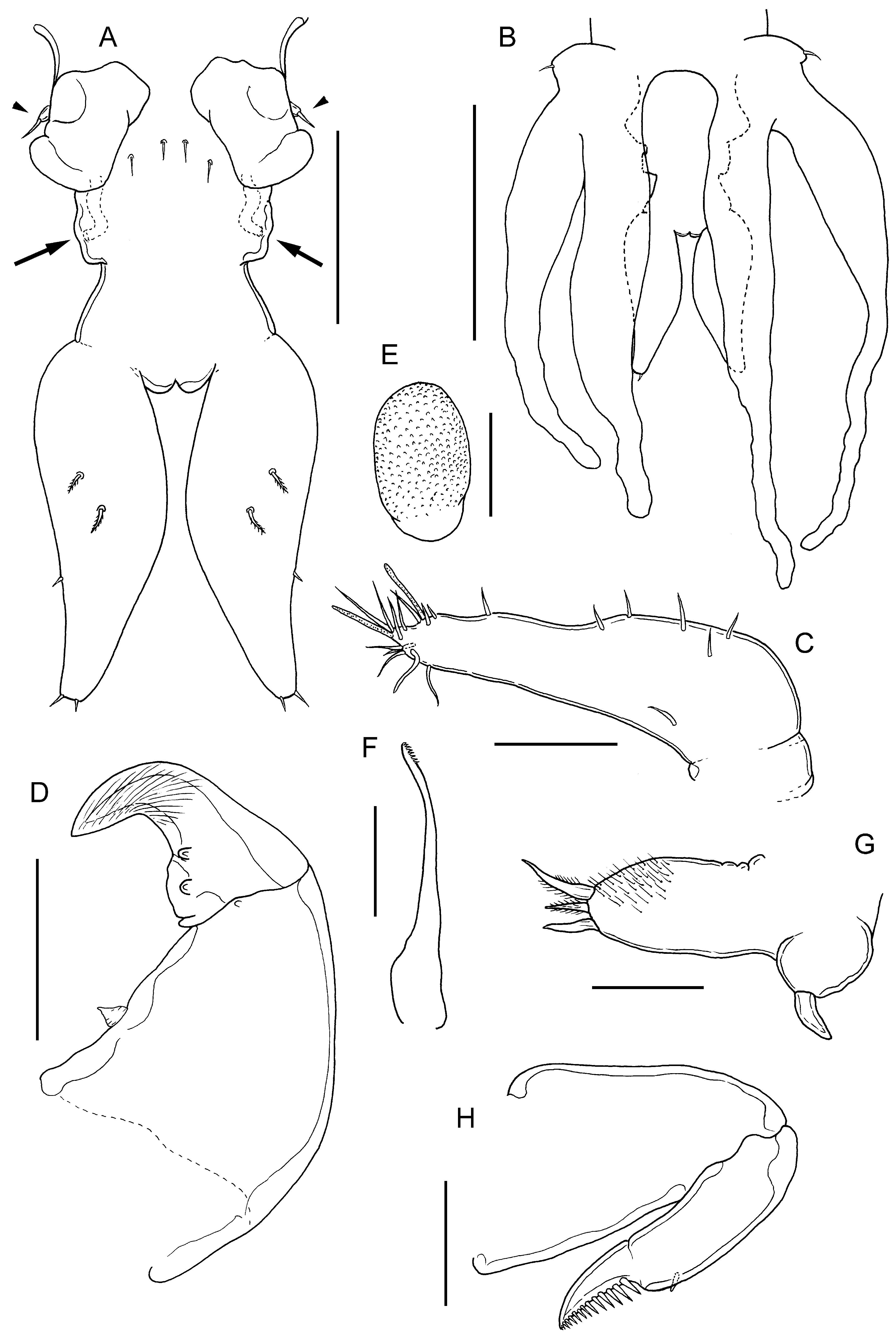
FIGURE 45. Sagum lativentris (Heller, 1865) n. comb., adult ♀. A, urosome, dorsal view showing vestigial fifth legs (arrow- heads), paired genital openings, copulatory pores (arrowed) and caudal rami; B, posterior end of trunk and urosome, ventral view showing extent of lobes of leg 4 relative to caudal rami; C, antennule; D, antenna; E, postantennal process; F, mandible; G, maxillule; H, maxilla. Scale bars A, 200 μm, B, 0.5 mm, C,E–H, 50 μm, D, 100 μm.
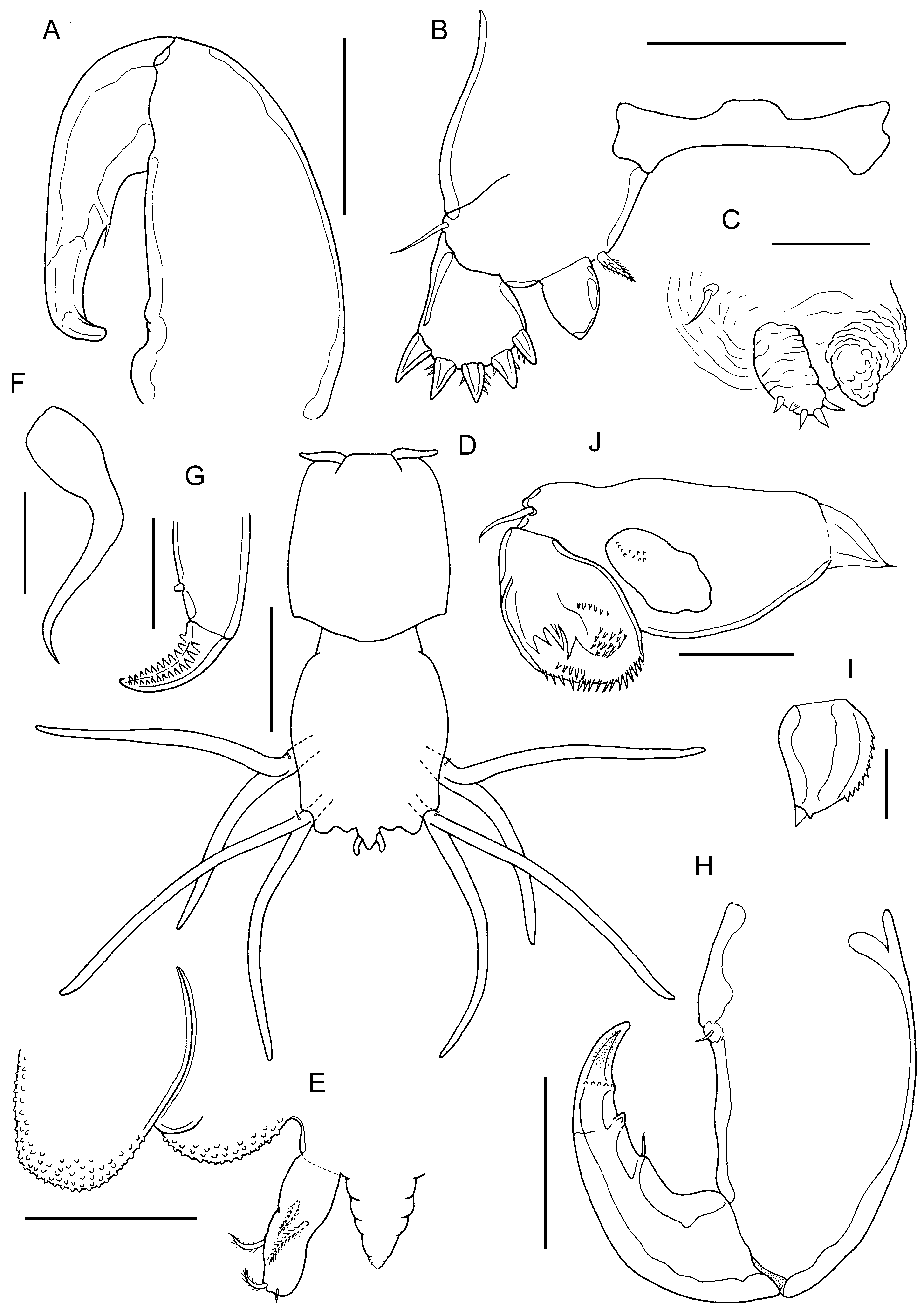
FIGURE 46. Sagum lativentris (Heller, 1865) n. comb., adult ♀. A, maxilliped; B, leg 1 and intercoxal sclerite; C, leg 2. Adult ♂. D, habitus, dorsal; E, genital operculum and caudal ramus on right side, ventral view; F, parabasal flagellum; G, tip of max- illa; H, maxilliped; I, endopod of leg 1; J, leg 2. Scale bars A,B,E,H, 100 μm, C,F,G,J, 50 μm, D, 0.5 mm, I, 25 μm.
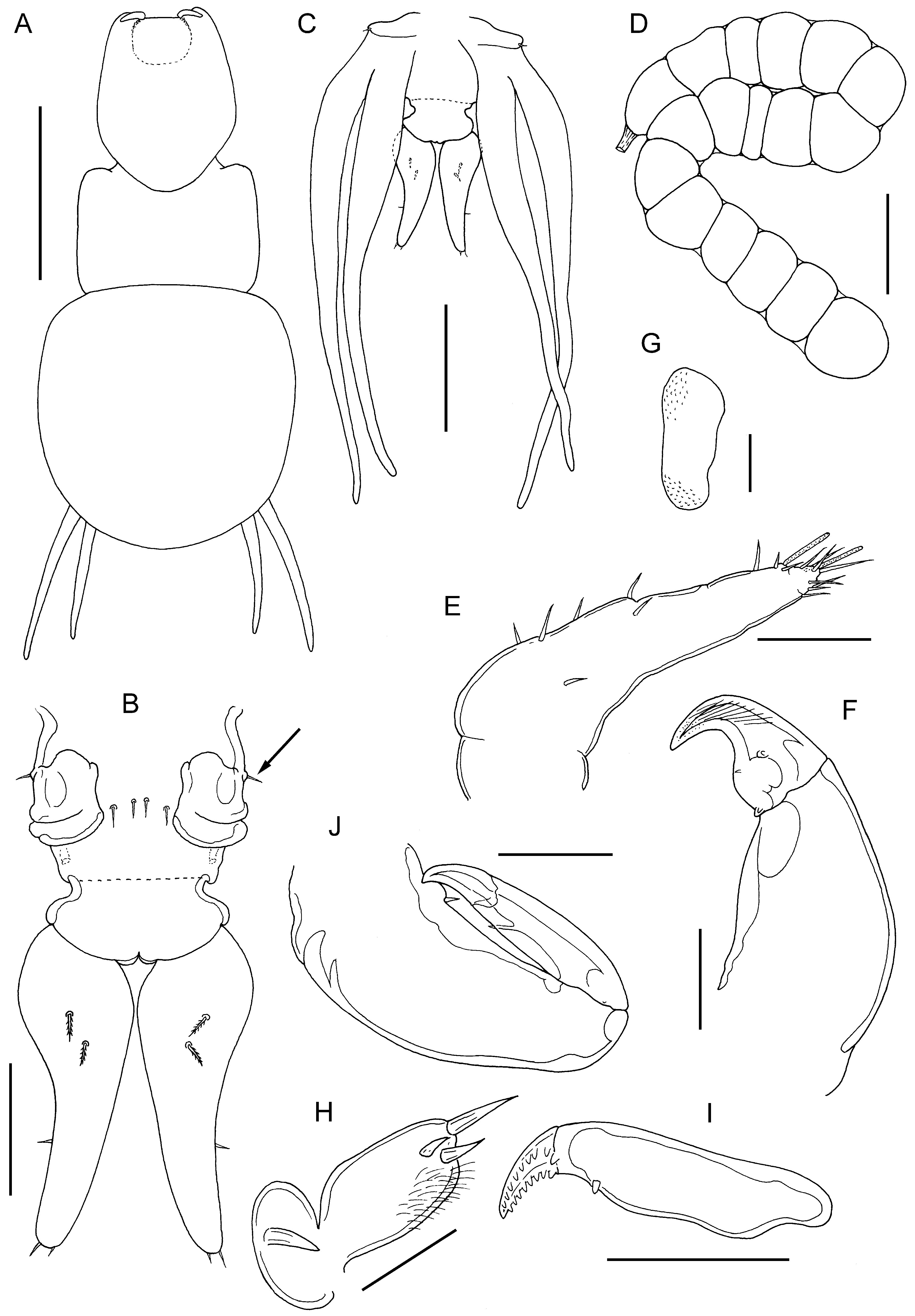
FIGURE 47. Sagum sanguineus (Song, in Song & Chen, 1976) n. comb., adult ♀. A, habitus, dorsal: B, urosome, dorsal view showing vestigial fifth legs (arrowed), paired genital openings and caudal rami; C, posterior end of trunk and urosome, ventral view showing extent of lobes of leg 4 relative to tips of caudal rami; D, egg sac; E, antennule; F, antenna; G, postantennal process; H, maxillule; I, basis of maxilla; J, maxilliped. Scale bars A, 1 mm, B,D, 200 μm, C, 0.5 mm, E,G,H, 50 μm, F,I,J, 100 μm.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Sagum lativentris ( Heller, 1865 )
| Boxshall, Geoff A., Bernot, James P., Barton, Diane P., Diggles, Ben K., Q-Y, Russell, Atkinson-Coyle, Toby & Hutson, Kate S. 2020 |
Sagum lativentris:
| Pattipeiluhu & Gill 1998 |
Lernanthropus lativentris
| : Pillai 1985 |
1 (by plazi, 2020-02-17 07:18:21)
2 (by ExternalLinkService, 2020-02-17 07:29:36)
3 (by ExternalLinkService, 2020-02-17 07:49:39)
4 (by ExternalLinkService, 2020-02-17 17:48:26)
5 (by veselin, 2020-02-27 10:06:36)
6 (by ExternalLinkService, 2021-10-20 03:17:43)
7 (by ExternalLinkService, 2021-10-20 05:35:24)
8 (by plazi, 2023-10-31 00:24:02)