
Cyphoderus mucrominimus, Oliveira & Alves & Zeppelini, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4350.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:22350FB0-A72A-41F5-A28F-159258BBC009 |
|
DOI |
https://doi.org/10.5281/zenodo.6017498 |
|
persistent identifier |
https://treatment.plazi.org/id/55468786-AC05-FFB7-5C85-C610B95A07EC |
|
treatment provided by |
Plazi |
|
scientific name |
Cyphoderus mucrominimus |
| status |
sp. nov. |
Cyphoderus mucrominimus sp. nov.
Figs 1–16 View FIGURES 1 − 7 View FIGURES 8 − 10 View FIGURES 11 − 15 View FIGURES 16 − 21. 16 − 17
Type material. Holotype, female on slide 7384, 01–19 III 2016. Brazil, Pará, Curionópolis, S 06°00’37.1”, W 49°38’14.8”, Serra Leste , cave, leg. SPELAYON team. Holotype deposited at CRFS-UEPB, João Pessoa, Paraíba GoogleMaps , Brazil. Paratypes: 3 females on slides, same data as holotype. labelled ref. # CRFS 7381 , # CRFS 7382 and the slide #3164 is deposited in MNUFRJ. GoogleMaps
Other material. See Table 1.
Description. Habitus typical of the genus, length 0.8 mm (average type specimen). Body color white. Eyes lacking, no ocular patch. Scales present on antennal segments I and II, head, body and furca (ventrally on manubrium, both sides on dens), and absent on legs and ventral tube.
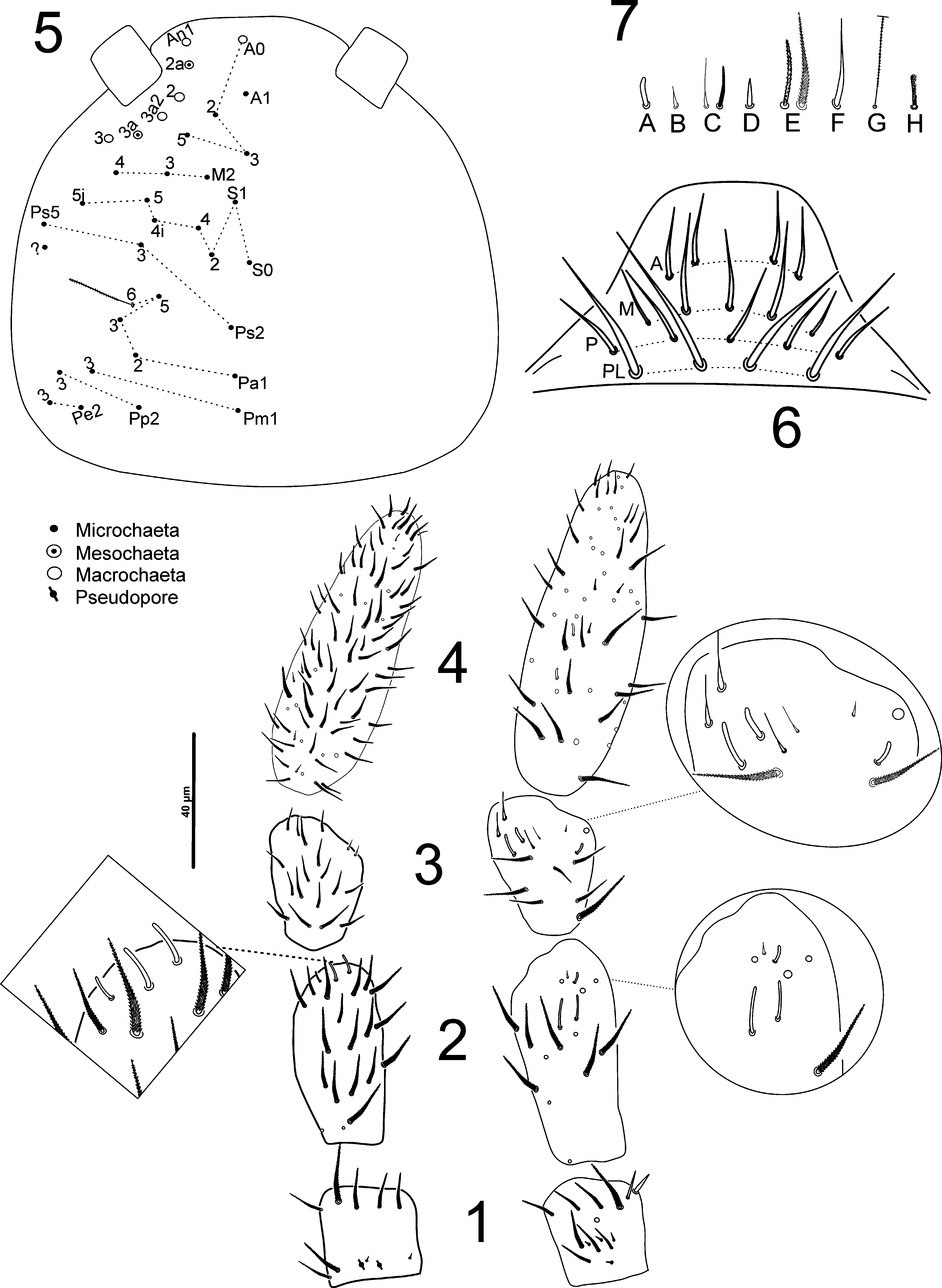
Antennae ( Figs. 1–4 View FIGURES 1 − 7 ) 1.35 times the length of the head, segments I: II: III: IV as 1: 2.3: 1.4: 3.4. Sensilla present on all antennal segments; Ant. I ventrally with four little sensilla ( Fig. 7B View FIGURES 1 − 7 ) at the base, a row with four apical ciliated chaetae, and two apical spine-like chaetae ( Fig. 7D View FIGURES 1 − 7 ) in right edge; dorsally with five distal chaetae, two proximal, two sensilla ( Fig. 7B View FIGURES 1 − 7 ) and two pseudopores at the base of antennal segment ( Fig. 1 View FIGURES 1 − 7 ). Ant. II ( Fig. 2 View FIGURES 1 − 7 ) ventrally and dorsally with three sense rods (per side) ( Fig. 7A View FIGURES 1 − 7 ) and plus one short apical sensillum ( Fig. 7B View FIGURES 1 − 7 ); AIIIO ( Fig. 3 View FIGURES 1 − 7 ), with 3 sense rods ( Fig. 7A View FIGURES 1 − 7 ), 4 bristle-like sensilla ( Fig. 7C View FIGURES 1 − 7 ) and 1 apical microsensillum ( Fig. 7B View FIGURES 1 − 7 ). Ant. IV ( Fig. 4 View FIGURES 1 − 7 ) apical bulb absent, dorsally with 4 basal sensilla ( Fig. 7B View FIGURES 1 − 7 ) and 3 distal bristle-like sensilla ( Fig. 7C View FIGURES 1 − 7 ); ventrally with 3 microsensilla ( Fig. 7B View FIGURES 1 − 7 ) more 2 sense rods ( Fig. 7A View FIGURES 1 − 7 ).
Head dorsal chaetotaxy ( Fig. 5 View FIGURES 1 − 7 ) with four macrochaetae and two mesochaetae at the An series; five chaetae at the A series (A0–A1–A2–A3–A5); M2–M4, S0–S1–S2–S4–S 4i –S5–S 5i, Ps2, Ps3 and Ps5 present in medioocellar to post-sutural areas; posterior chaetae Pa1–Pa2–Pa3, Pa5–Pa6 (bothriotrix), Pm1, Pm3, Pp2, Pp3, Pe2 and Pe3 present. Labrum with amp rows of chaetae as 4,5,5 smooth chaetae with 4 prelabral chaetae larger and longer than others ( Fig. 6 View FIGURES 1 − 7 ). Labial palps with all papillae present ( Fig. 8 View FIGURES 8 − 10 ), chaetae a1, b1–4, d1–4, e1–3, e6 and lateralprocess (lp) present. Labial triangle formula M1 M2 R E L1–2 A1–4 ( Fig. 9 View FIGURES 8 − 10 ). Chaetae along ventral groove ciliated with one microsensillum laterally ( Fig. 9 View FIGURES 8 − 10 ).
Main macrochaetae on segments Th. II to Abd. IV as 00/0226+14. Body chaetotaxy (as in Fig. 10 View FIGURES 8 − 10 ), anterior hood of Th. II with a collar of two or three rows of ciliated and capitated macrochaetae (not represented in the drawing). Th. II with a2, m1–2, m4–6, p1–4, p6, p6e and one antero-lateral microchaeta. Th. III with m2, m7, m6p, p2–4, p6 and one microchaeta as. Abd. I with five chaetae m2–6 and as. Abd. II with two bothriotricha a5 and m2 each surrounded by 2 fan-shaped chaetae each. m3, m5 macrochaetae, m3e, a6, m6–7, p6–7 and as present. Abd. III with three bothriotricha a5, m2 and m5 surrounded by 3,3,2 fan-shaped chaetae respectively; chaetae a6–7, pm6, am6, p6–7 present, microchaeta as present and one extra sensillum near p6 chaetae. Abd. IV with three bothriotricha T2, T4 and E4 surrounded by 4,1,2 fan-shaped chaetae respectively; A4 and A5 with accessory sensillum. A3, A6, B3–6, C1, C1p, C2–4, T3, T5, T7, D1, D1p, D2, D3, De3, E1–4, E4p, Ee12, F1–3 and Fe2–4 present, posterior row of chaetae at the distal edge of Abd. IV with 7+7 mesochaetae, 2+2 pseudopores present on the posterior edge of the segment.
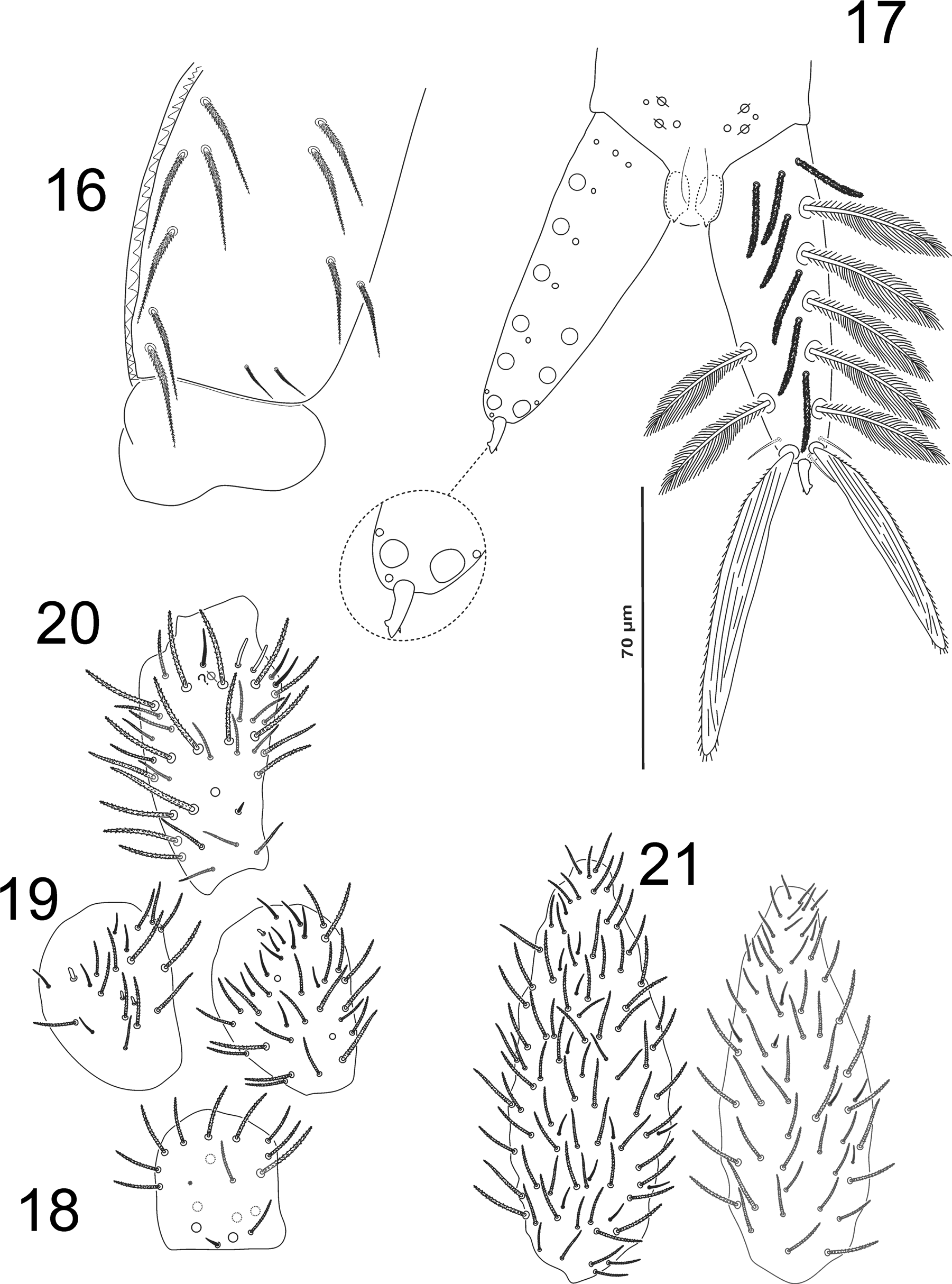
Legs ( Figs. 11–15 View FIGURES 11 − 15 ) without scales, trochanteral organ V-shaped with 9 ciliated chaetae ( Fig. 13 View FIGURES 11 − 15 ). Tibiotarsi with tenent hair apically clavated ( Fig. 15 View FIGURES 11 − 15 ). Claws broad, with one tooth in external lamella; a pair of inner basal teeth of unequal size and one unpaired subapical tooth ( Fig. 15 View FIGURES 11 − 15 ). Unguiculi pointed and broad ( Fig. 15 View FIGURES 11 − 15 ).
Ventral tube without scales, anteriorly with 6+6 macrochaetae, 2+2 distal mesochaetae and with 4+4 macrochaetae in posterior side, all ciliated ( Fig. 16 View FIGURES 16 − 21. 16 − 17 ).
Furcula ( Fig. 17 View FIGURES 16 − 21. 16 − 17 ). Manubrium 1.4 times as long as dens; Manubrial plate with 2 ciliate chaetae and 2 pseudopores. Dens 8 times longer than mucro, dorsally with 2 rows of feathered scales, 6 external and 3 internal, and 4 ciliated between the two rows; proximal outer part of dens with 3 ciliated chaetae and 3 smooth apical chaetae on ventral side, 2 at the base of external scale and one at the base of internal scale. Mucro vestigial (9,5µm), bidentate apically, with one small subapical tooth (aAI).
Etymology. The species is named after the reduced size of mucro.
Distribution. The species was recorded from Amazon Rainforest iron caves at Curionópolis (Serra Leste) and Parauapebas, Pará State, northern Brazil.
Ecology. Collected in four iron caves in Serra Leste and five iron caves in Parauapebas. These caves are inserted in the Amazon forest, mostly small caves whose conditions vary according to the surface weather. The altitude is between 500– 670m. The climate according to Köppen’s system is Af: Equatorial rainforest, fully humid ( Köppen 1936, Kottek et al. 2006).
Remarks. C. mucrominimus sp. nov. differs from Brazilian tridenticulati, Cyphoderus arlei in the reduced size of mucro which is less than 1/5 the length of the dens. They also differ in the shape of the chaetae r in labial triangle, which is a normal seta in the new species and is reduced in C. arlei , the TO with 9 chaeta (versus 12 in C. arlei ), dens with 6 external and 3 internal feathered scales (versus 6~7 external and 5~7 internal feathered scales in C. arlei ).
The new species resembles somehow those of the genus Pseudocyphoderus Imms, 1912, mainly in the shape and size of the mucro. Nevertheless, the new species clearly belongs to the genus Cyphoderus by presenting fully developed mouth parts, while the species of Pseudocyphoderus are known to present a reduction of the pars incisiva on the mandible and a rather hypognath position ( Delamare-Debouteville 1948). Mari-Mutt (1977) described a species (P. mellitophilous Mari-Mutt 1977) with well-developed mouth parts and lacking mucro as in C. mucrominimus sp. nov., nevertheless, P. mellitophilous presents a set of morphological features, adaptive modifications and a large body size, which clearly differs from the new species, as examples body length over 3 mm, dorsoventrally flattened body, “Lepismiform” aspect, large trochanters, modified metathoracic legs and large tenent hairs. Furthermore, the ventral tube chaetotaxy strikingly differs from that of Cyphoderus ( Yoshii 1987) .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |