
Trimma hoesei Winterbottom, 1984
|
publication ID |
https://doi.org/10.11646/zootaxa.3934.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:11C2A2CB-30B3-4694-B379-AE9D47332F0C |
|
DOI |
https://doi.org/10.5281/zenodo.5621512 |
|
persistent identifier |
https://treatment.plazi.org/id/5519879A-B822-F366-FF1F-FDBE6F76EE2F |
|
treatment provided by |
Plazi |
|
scientific name |
Trimma hoesei Winterbottom, 1984 |
| status |
|
Trimma hoesei Winterbottom, 1984 View in CoL
Forktail Pygmygoby
Figs. 1 View FIGURE 1 B, 15–16, Pl. 1 H
Trimma hoesei Winterbottom, 1984:704 View in CoL (Chagos Archipelago, central Indian ocean); Allen et al., 2004:331; Allen & Erdmann, 2012:940 (Cocos (Keeling) Ids to Fiji).
Trimma View in CoL sp. 4: Kuiter, 1998:204 ( Maldives).
Trimma View in CoL sp. 11: Kuiter & Tonozuka, 2004:706 ( Indonesia).
Material. The type material ( 128 specimens, 9.0– 24.2 mm SL) from the Chagos Archipelago, plus the following:
Australian material. Great Barrier Reef: One Tree Id: AMS I.20577-008, (23), 30 m. Cocos (Keeling) Ids: WAM P. 29906.017, (15), 20– 31 m.
Other Material. Indonesia: Bone Rate Id: BPBM. Moluccas: BPBM. Raja Ampat Ids: ROM 85205, 4(20–26), 18–23 m; ROM 84892, (29), 56 m; ROM 85331, 11(11–22), 66 m; ROM 85342, 27(18–28), 20– 27 m. WAM P. 32925.005, (24), 40– 50 m. Palau: Koror Id: AMS; ROM 74773, 2(24.1–26.0), 15.2–26.5 m; ROM 80356, (23.3), 73 m; ROM 80366, 8(18.6–27.1), 17.4 m; ROM 80394, (21.0), 19.8–27.4 m. Helen Reef: ROM 83276, 25(12.6–26.2), 22–34 m; ROM 83303, 60(21.1–23.6), 27m. New Britain: WAM P. 28178.012, 2 (16–21), 33– 40 m. Papua New Guinea: Manus Id: WAM P. 27826.068, 7 (19–20), 35–41 m; WAM P. 27826.093, 6 (18–20), 35–41 m; WAM P. 28178.012, 2 (16–21), 33– 40 m. Ninigo Id: USNM 313556, 0–4.6 m. Philippines: Balicasag Id: USNM 313544, 6(15–24), 0– 24 m. Southern Cebu: USNM 263539, 13(15–22), 18–24 m; USNM 333312, (18), 18– 24 m. Solomon Ids: Florida Id: AMS I.17500–027, (20), 20– 30 m.
Diagnosis. A species of Trimma with the posterior one quarter or less of the predorsal midline scaled, these scales usually overlapping rows from either side rather than centred on the midline; a relatively broad bony interorbital (about pupil diameter in width); 14–15 pectoral-fin rays with the upper three and lower 2–3 unbranched; multiple, sequential branches to the first four pelvic-fin rays (up to five branch points, six branch tips); 3–4 + 14–15 = 17–19 gill rakers on the outer face of the first gill arch; a forked caudal fin ( Fig. 16 View FIGURE 16 ); ventral half of body translucent yellow-orange to red in life, pale above; body uniformly straw-coloured in preservative with darker diffuse saddles across the dorsum.
Description. The following description is taken mostly from Winterbottom (1984), with corrections made for differences in methodology, omissions and errors in text and figure derived from 10 (of 60) specimens from Helen Reef, Palau (ROM 83303, 10(21.1–23.6), because of the paucity of Australian material of this species.
Dorsal fin VI + I 9–10 (usually 10), second and third spines longest with second reaching between the base of third ray of second dorsal fin and anterior region of the peduncle, third spine may reach posteriorly as far as base of fourth fin ray, first ray of second dorsal fin usually branched, last few rays increasingly elongated so that last reaches posteriorly to, or almost to, dorsal procurrent caudal-fin rays, last ray consisting of two unbranched rami; anal fin I 9–11 (usually 10), first ray branched, last few rays increasingly elongated so that last reaches to, or almost to, ventral procurrent caudal-fin rays, anterior ramus of last ray branched, posterior ramus unbranched; pectoral fin 14–16 (usually 15), with three dorsal and 2–3 ventral rays unbranched, others branched once dichotomously; pelvic fin I 5, fifth ray branched once dichotomously reaching posteriorly to urogenital papilla and 78–92% of fourth, which reaches posteriorly to region of first few elements of anal fin, rays 1–4 with 3–5 sequential branches (i.e. 4–6 branch tips per ray), no fraenum, basal membrane 7–12% fourth ray; caudal fin forked. Scales on sides of nape extend to above middle of opercle. Lateral scales 23–24; transverse scales 8; predorsal scales 1–5 ( 1–3 in the Palau material), count includes scales from left and right sides that overlap in the midline, confined to area just in front of origin of first dorsal fin; body scales ctenoid; scales anterior to a line between pectoral-fin base and first dorsal-fin origin cycloid or weakly ctenoid; scales in belly midline cycloid anteriorly becoming ctenoid posteriorly; pectoralfin base with 4–5 vertical rows of cycloid scales; 5–6 scales in irregular rows across midline anterior to pelvic fin; no scales on cheek or opercle. Outer row of upper jaw and inner and outer rows of lower jaw teeth consisting of curved, evenly spaced, enlarged canines, other teeth short, conical and in irregular rows. Tongue broadly to moderately pointed, three-quarters pupil diameter in width. Gill opening extending anteroventrally to below midpupil; first gill arch with 3–4 + 14–15 outer rakers. Anterior nares in a short, slim tube; posterior opening pore-like with a slightly raised rim; nasal sac slightly elevated; nasal apparatus confined to middle half of snout. Interorbital broad (~ pupil diameter), with two shallow troughs on either side of a median ridge, interorbital region forming a flattened, rounded 'W' in cross section ( Fig. 1 View FIGURE 1 B); epaxialis reaching anteriorly to a point perpendicular with middle of pupil, a dermal ridge in dorsal midline extending posteriorly to near anterior extent of predorsal scales. Abdominal/caudal vertebral transition Type A.
Colour pattern. Freshly collected. A 24.1 mm SL female from Helen Reef, Palau ( Fig. 16 View FIGURE 16 ) with translucent dorsum, scale pockets thinly outlined in red and scattered, half pupil diameter yellow spots, usually below posterior margins of scales. Ventrum red, margined distally by diffuse, half pupil-diameter wide orange stripe just lateral to anal fin origin. Neural canal visible through body as diffuse dark grey stripe; dorsal surface of swimbladder visible as series of diffuse charcoal blotches. Ventral part of head off-white, cheek with scattered red chromatophores and diffuse, light yellow, oblique stripe, and opercle red with continuation of yellow cheek stripe. Snout reddish, with oblique yellow stripe from midregion of anterior margin of eye across upper jaw; anterior halves of both jaws red. Iris golden, margined dorsally and ventrally by dark purple stripe with similar stripe through middle. Nape with scattered red chromatophores and diffuse yellowish spots and blotches. Elements of dorsal fins red, accentuated at base with vague spots, and narrow (one-quarter pupil width) translucent proximal stripe with scattered red melanophores followed by diffuse, half-pupil width yellow stripe. Distally, first dorsal fin membranes translucent with scattered iridocytes and dark chromatophores, second dorsal fin similar, but with occasional diffuse yellow spots centred on fin rays and dark chromatophores concentrated along margin of fin. Anal fin yellow with half pupil diameter red basal stripe and dark chromatophores concentrated along distal edge. Caudal fin reddish, with four yellow stripes, two along rays forming elongated lobes of fin, and two arising from common source at middle base of fin over ural centrum, diverging and widening posteriorly. Distal margins of hypural outlined by thin red bar, interrupted at midline by yellow stripe. Pelvic fins hyaline with scattering of yellow and grey chromatophores. Freshly collected, 21.3 mm SL female from Chagos (see Winterbottom, 1984, fig. 4) and two other photographed specimens from Palau (23.3 and 26.0 mm SL females) very similar. The latter have scale pockets on dorsum redbrown or grey; and caudal fin reddish-purple with faint, diffuse yellow stripes, outer pair (along fin rays forming upper and lower lobes of fin) about three times wider than inner pair. Live: specimen photographed in Maldives by R. Kuiter. Translucent with red wash extending from abdomen to just behind eye, four light blotches on otherwise black nerve cord, several irregularly spaced blue dashes on head (including top of eye), and white tip to elongated second dorsal spine. Pair photographed spawning on alcyonarian by G. Allen at East Cape, Milne Bay, PNG in about 15 m: pale cerise ventrally, with yellow nerve cord, dorsum semi-translucent purple-red. Several specimens from Palau photographed by P. Colin (see Pl. 1 H): similar to Maldives specimen, but translucent dorsum tinged with red or yellow, some show diffuse or well demarcated pale stripe in middle of snout.
Preserved. Pale straw-yellow with an irregular, diffuse and variably-developed line of chromatophores in midline of snout and anterior interorbital region, continuous line of melanophores in midventral septum between base of last anal-fin ray and ventral procurrent caudal-fin rays; anal fin dusky, few scale pockets outlined with dark chromatophores dorsally on body, forming series of diffuse saddles.
Etymology. Named for Douglass F. Hoese, co-author of this paper, in recognition of his many contributions to gobioid systematics.
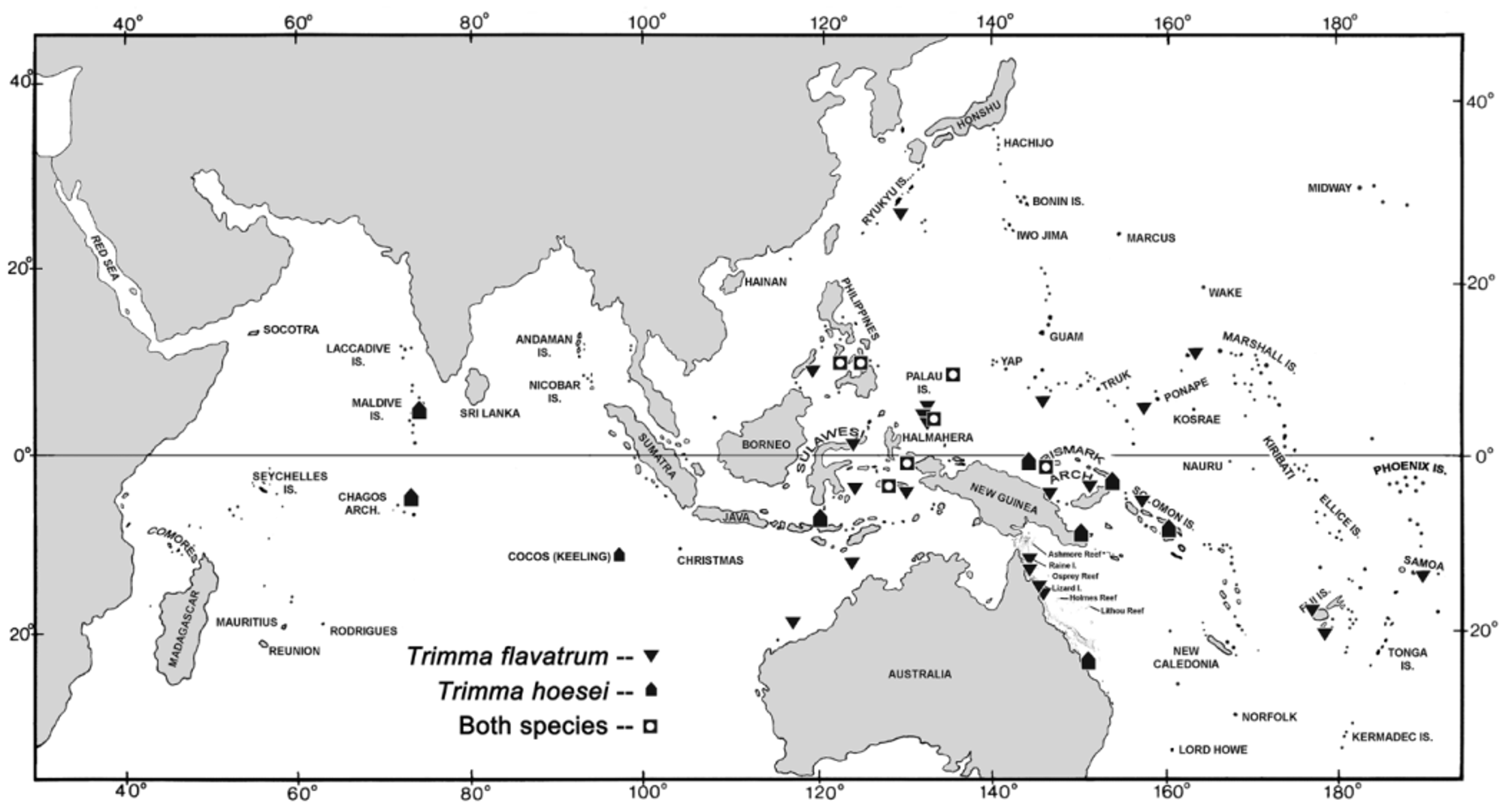
Distribution. The species has been collected at Cocos (Keeling) Ids, and photographed at the Maldives, the only other Indian Ocean localities beside the type locality at the Chagos Archipelago. We have examined preserved material from off Timor and the Moluccas, the Philippines, Palau, Raja Ampat, Papua New Guinea, New Britain and the Solomon Islands in the western Pacific ( Fig. 15 View FIGURE 15 ).
Comparisons. Trimma hoesei is unique among members of the genus in possessing a forked caudal fin ( Fig. 16 View FIGURE 16 ). It differs from all other species of Trimma with a broad, bony interorbital except T. taylori in having the first four pelvic-fin rays with 3–4 sequential branches (as opposed to a single sequential branch). It differs further from T. taylori in that the second and third dorsal spines are elongated in both sexes (elongated only in males of T. taylori ), and in having only a single expanded haemal arch (vs. 4–5 expanded haemal arches—see Winterbottom, 1984, fig. 26). See Comparisons section of T. taylori for further differences.
Four specimens (Helen Reef, Palau, n = 2 and Raja Ampat, Indonesia, n = 2) were sampled for CO1, with only a minor difference between them ( Winterbottom et al., 2014). One of the Helen Reef specimens was phenetically closer to the two Raja Ampat specimens than to the other Helen Reef specimen.
Discussion. Winterbottom (1984:704) stated the basal membrane of the pelvic fin was “... delicate, apparently about one-third of the length of the 5th ray”. The material from Palau (ROM 83303) is in very good condition and allows a quantification of the membrane. It is ‘U’-shaped, with the apex directed anteriorly in the midline and 7–12% of the length of the fourth ray. The arms of the U taper towards the fifth ray, and attach to it at about onethird of the length of the ray.
This species has been collected infrequently, and the largest lots came from well-developed caves in a vertical wall at about 40 m in the Chagos Archipelago and from similar caves at 27m at Helen Reef, Palau. G. R. Allen provided us with a photograph of a pair of T. hoesei spawning on an alcyonarian coral that was taken in Milne Bay, PNG at a depth of 15 m. The species is probably more widely distributed in the Indo-west Pacific (sensu Winterbottom et al., 1989) than current records indicate.
Trimma hoesei is normally found on drop-offs in depths of 20– 50 m. In Australia, it is known only from single specimens from the southern Great Barrier Reef and from Cocos (Keeling) Islands.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Trimma hoesei Winterbottom, 1984
| Winterbottom, Richard & Hoese, Douglass F. 2015 |
Trimma
| Kuiter 2004: 706 |
Trimma
| Kuiter 1998: 204 |
Trimma hoesei
| Allen 2012: 940 |
| Allen 2004: 331 |
| Winterbottom 1984: 704 |