
Agonochaetia Povolný, 1965
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4300.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:EEAD66E3-68E2-488C-A4E1-87D145E880C8 |
|
DOI |
https://doi.org/10.5281/zenodo.6033117 |
|
persistent identifier |
https://treatment.plazi.org/id/541787FC-FF8D-AA41-7A96-20F3D7ACF898 |
|
treatment provided by |
Plazi |
|
scientific name |
Agonochaetia Povolný, 1965 |
| status |
|
Genus Agonochaetia Povolný, 1965 View in CoL
Agonochaetia Povolný, 1965: 487 View in CoL . Type species: Agonochaetia incredibilis Povolný, 1965: 487 View in CoL , fig.10, by original designation. Huemer & Karsholt 2010: 294.
= Sautereopsis Povolný, 1965: 488 View in CoL . Type species: Gelechia terrestrella Zeller, 1872: 111 View in CoL , by original designation. Synonymized by Sattler, 1968: 119.
Diagnosis. Agonochaetia is characterized by the following unique combination of characters in the male genitalia: 1) a pair of needle-like glandiductors that are as long or longer than the phallus, outwardly and upwardly curved, slightly sinuate, with their anterior bulbous base closed (blind sac), and free from each other and from the phallus; 2) mesio-distal process of the gnathos absent; 3) sacculus large and heavy, inwardly bent at 90 degrees, commashaped. In the female genitalia (known for only four of the nine species) the antrum is elongate, anteriorly tapered, funnel-like, as long or longer than sternum 8, and the signum is an elongate, triangular base plate with a straight spine. Externally Agonochaetia moths are rather large, broad-winged, mostly unicolorous brown Gnorimoschemini which may be confused with Lutilabria species, and also with some Filatima , Chionodes , or other large and indistinctly marked or unmarked brown gelechiids with broad wings.
Description. Adult ( Figs. 1–12 View FIGURES 1 – 12 ). Rather large (wingspan 13–21 mm), broad-winged Gnorimoschemini with mostly unicolorous brown or beige brown forewings, in some species with 2 or 3 black spots in discal and postmedian area and along termen. Labial palpus slender, segment 3 shorter than 2. Antenna with flagellum filiform. Forewing and hindwing broad, ratio width/length = 0.26–0.30 in FW, 0.30–0.36 in HW. Hindwing sinuation weakly suggested or indistinct.
Wing venation ( Fig. 15 View FIGURES 13 – 16 ). Forewing with R1–R5 running to costa; R4 and R5 stalked, with M1 extended from their common stalk; pterostigma absent; in some specimens an accessory cell present between R1 and R2 (observed in A. intermedia ); upper portion of cell open; CuP absent. Hindwing with R1 running into Sc beyond base of wing; M1 extended from Rs beyond cell closure; upper portion of cell open, lower portion with closure perpendicular with long axis of wing; M3 and CuA1 stalked (the long stalk in Fig. 15 View FIGURES 13 – 16 is probably atypical; in another A. intermedia specimen in which the veins were examined without a cleared preparation, the M3-CuA1 stalk was short on the right side but long on the left; in the specimen illustrated, the stalk was short on both hindwings).
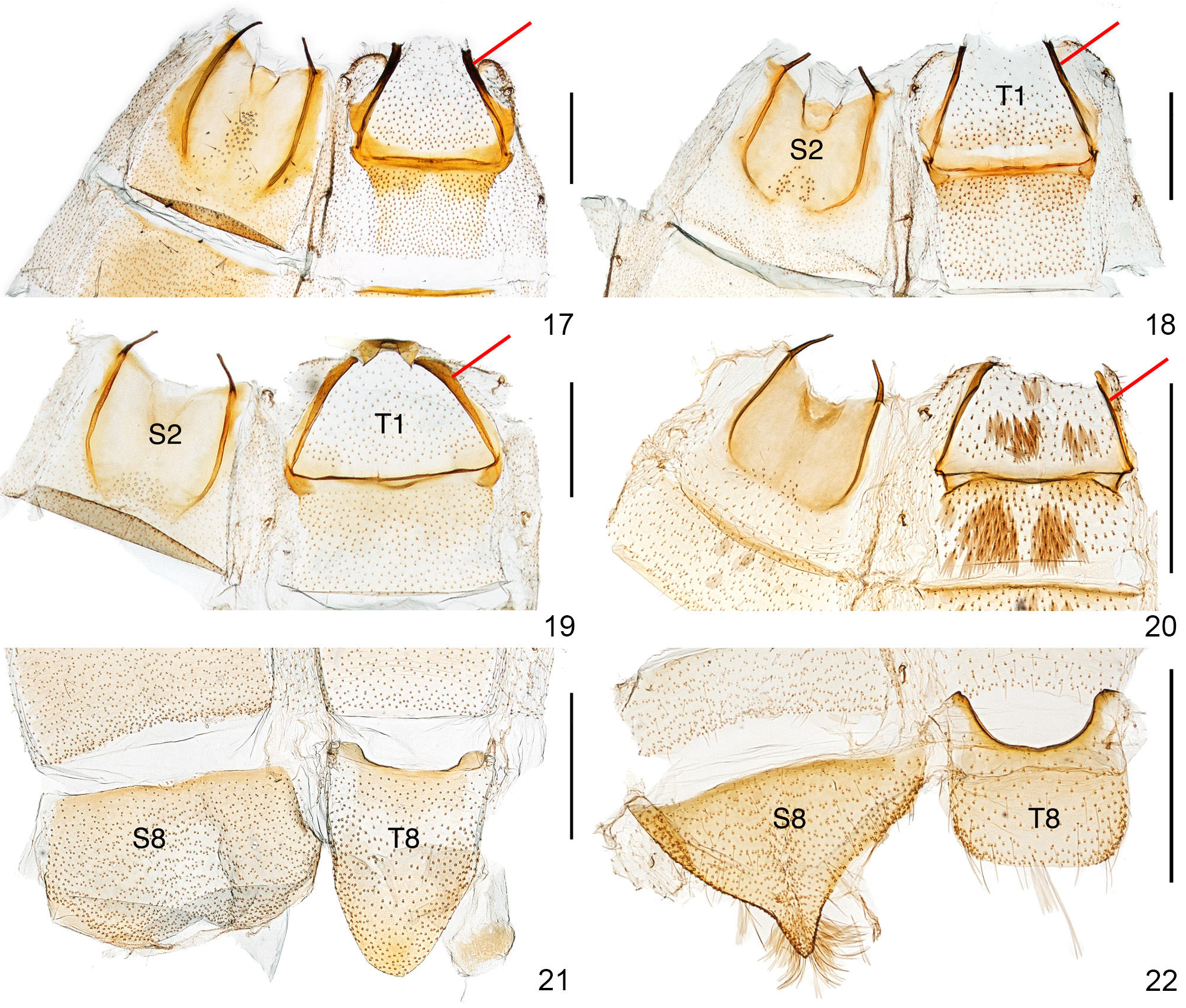
Abdomen ( Figs. 19, 21 View FIGURES 17 – 22 ). Lateral ridges of tergum I (so-called marginosclerites; see Kristensen 2003: 94) unmelanized throughout. Tergum VIII tongue-shaped, anterior margin thickened. Sternum VIII transversely subrectangular or sub-trapezoid, anterior margin thickened, posterior margin slighty indented; in male, without coremata.
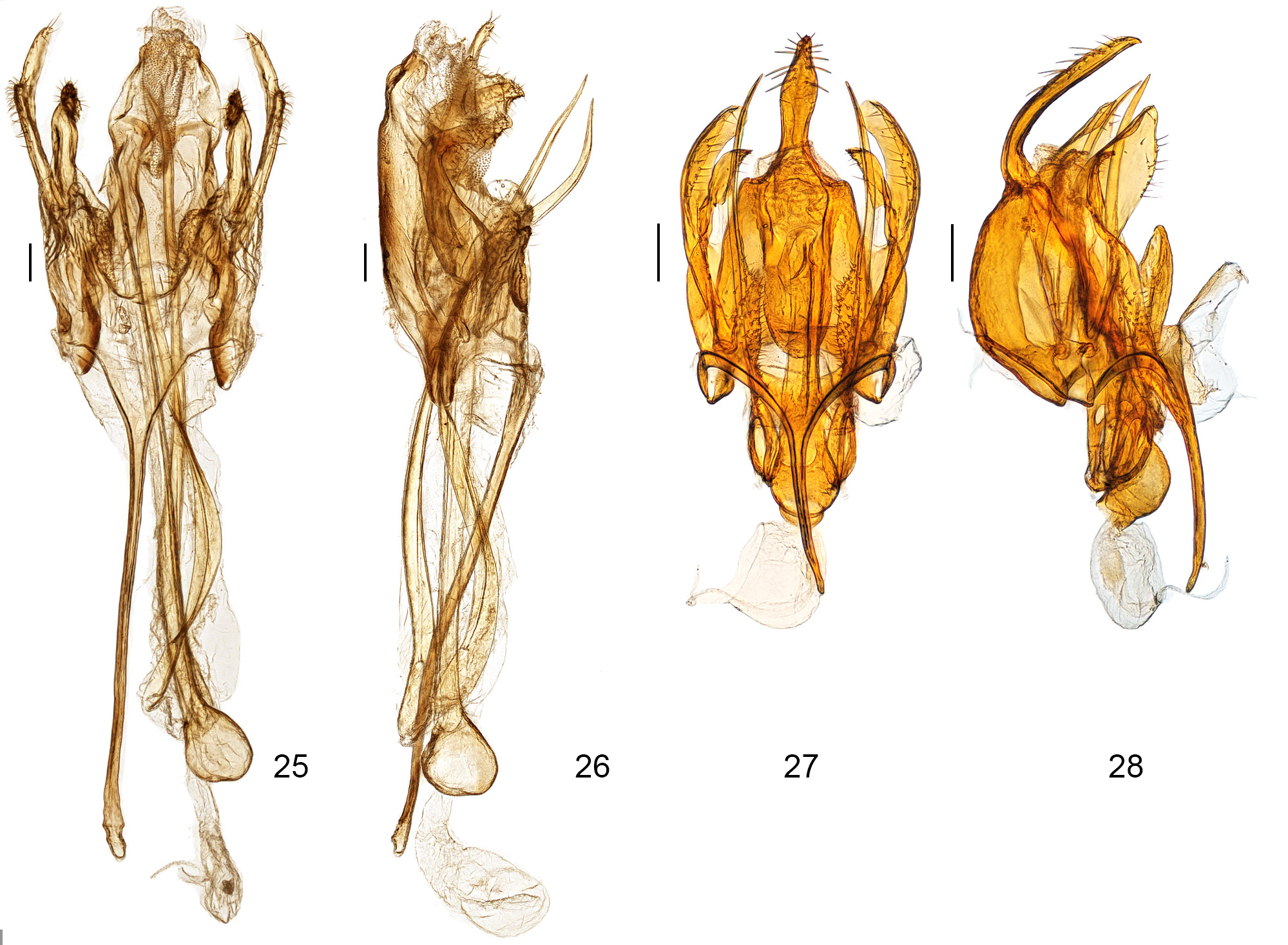
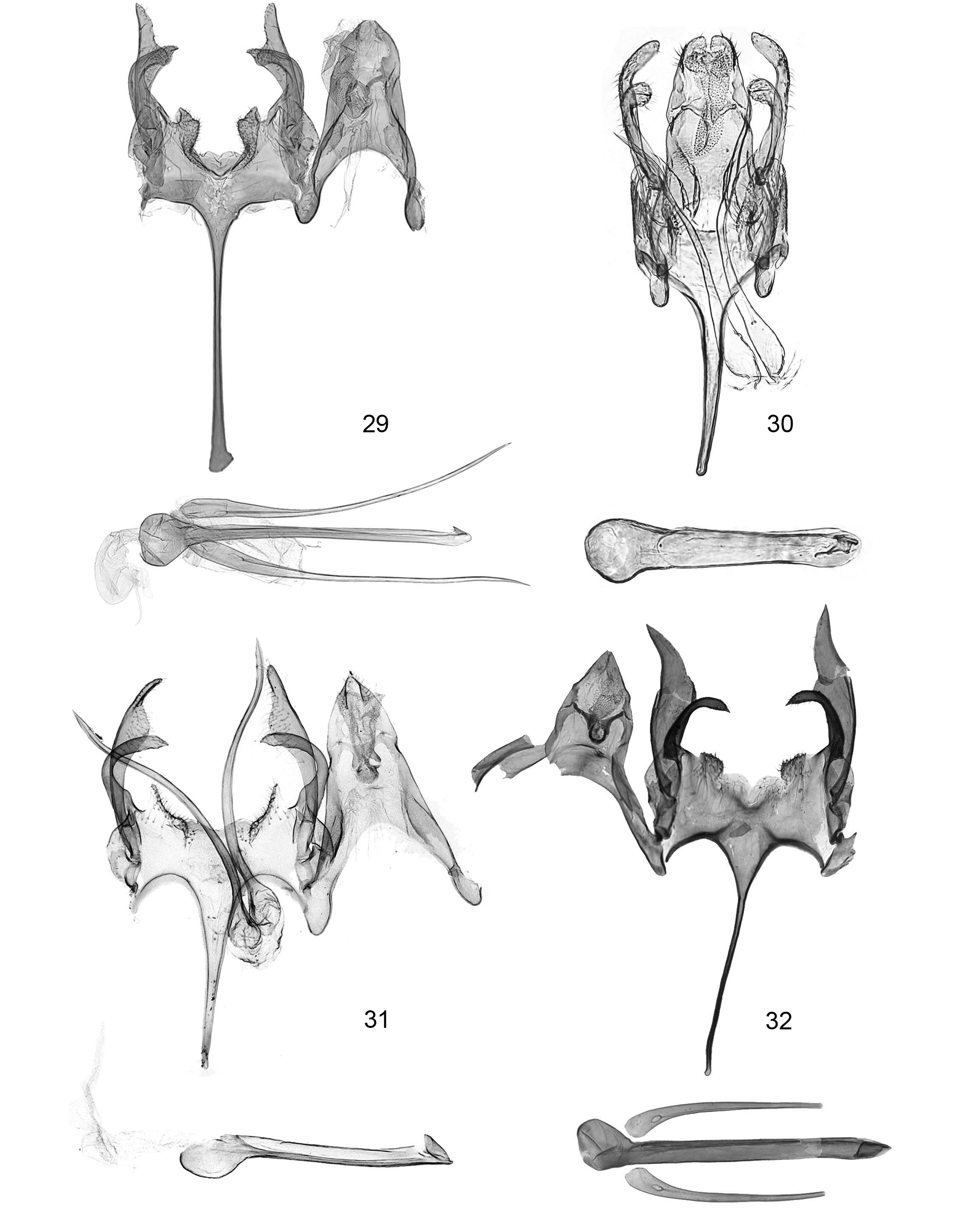
Male genitalia ( Figs. 25–26 View FIGURES 25 – 28 , 29–35 View FIGURES 29 – 32 View FIGURES 33 – 36 ). Tegumen triangularly elongate with deep and roundly arched anterior notch (TGN/TGL = 0.40–0.67; ratio may vary depending on amount of flattening). Uncus trapezoid, 0.3–0.4x length of tegumen, anterior margin fused to distal margin of tegumen and with delineation from latter somewhat indistinct as a dorso-lateral sulcus/impression, apical margin transversely straight, lined with several short setae, with shallow medial indentation. Gnathos with proximal arms short, stubby, apices club-like or hatchet-like and not mesially joined; mesial process vestigial, reduced to small, thin, bracelet-like sclerotized band embedded in apical part of culcitula and free from proximal arms. Culcitula a large, elongate, pouch-like lobe extended from apex of uncus to slightly below apex of proximal arms of gnathos. Vinculum transverse, with a deep V-shaped medioposterior emargination, ventral wall between cucullus base and vincular lobes finely wrinkled; vincular lobes hump-like, finger-like, pointed or squarish, mesio-dorsal surface densely and finely setose; saccus very long and thin, broadened basally, 0.85– 1.6x length of valva. Cucullus extended beyond apex of uncus, narrow, inner (ventral) margin variously widened or lobate in distal third, distal ¼ slender, digitiform or pointed. Sacculus strongly downcurved at almost 90° (appearing incurved in flattened preparation), apex rounded or widened with shallow concavity and acuminate tip. Paired glandiductor lobes needle-like, outwardly and upwardly curved, slightly sinuate, as long or longer than phallus, anterior portion variously dilated; lobes free from each other and from phallus, situated on each side and slightly dorsad of phallus at end of deep inpocketing of diaphragma. Phallus straight, slightly longer than saccus, apex with triangular dorsal tooth, caecum markedly bulbous, inception of ductus ejaculatorius dorso-anterior on caecum.
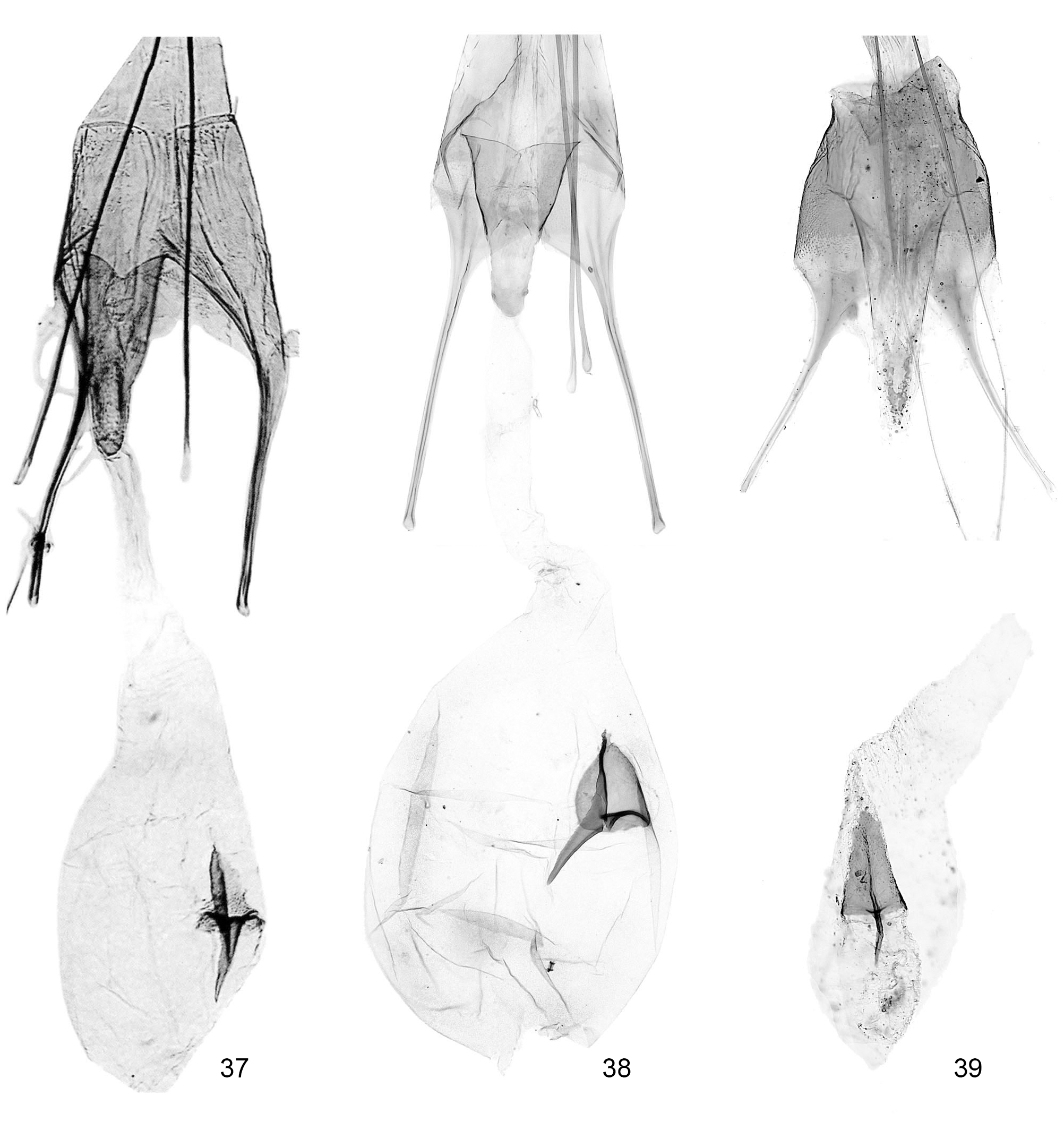
Female genitalia ( Figs. 37–41 View FIGURES 37 – 39 View FIGURES 40 – 42 ). Segment VIII longer than broad, without distinct sclerotized modifications, only short longitudinal folds ventrally. Anterior apophyses longer than segment VIII (AA/S8L = 1.5–3.5), base gradually widened and merged with wall of segment VIII. Antrum elongate, funnel-shaped, with posterior margin variously concave or indented. Ductus bursae membranous throughout, without ornamentation, sclerotization or spicules; corpus bursae ovoid to oblong; signum with sub-triangular basal plate and straight, thin, hook.
Biology. Host plants and immature stages unknown. Huemer & Karsholt (2010) speculated that larval hosts possibly include Caryophyllaceae based on its putative relationship to other Gnorimoschemine genera with Caryophyllaceae as host plants [‘caryocoloid’ Gnorimoschemini ( Povolný & Šustek 1988)]. Povolný (1990) speculated that some species of Agonochaetia might be oligophagous (as in Sattleria dzieduszyckii Nowicki, 1864 ) without explaining the grounds for such speculation. All Palearctic species inhabit xeric steppe-slopes, in Europe at elevations up to 2000 m. Even less is known about the Nearctic species: no host or habitat information is known for the western A. conspersa which was found at a few scattered localities in the West, whereas the new species described below was found, unexpectedly, in a suburban backyard in Eastern Canada.
Distribution. Holarctic, with seven described species recorded from the Swiss Alps (Graubünden: Münstertal, Bergun, Ramosch; Wallis: Zermatt), Austrian Alps (Fliess, Kauns), Balkan ( Bulgaria, Romania) Uzbekistan (Margellan), southern part of European Russia (southern Ural Mts. and Volgograd), southern Siberia (Altai and Tuva Mts., Buryatia, Zabaikalskiy krai), the Russian Far East (Khabarovsk krai), western Kazakhstan, Japan (Hokkaido), and two from North America (one in the East, one in the West).
Remarks. The species of Agonochaetia seem to be generally rather rare, xeromontane taxa with a scattered (islet-like) distributional pattern. The scantiness of material is a major hindrance for the validation of taxa in this genus ( Huemer & Karsholt 2010) as well as for assessing species distributions.
The fact that a junior synonym of the European A. terrestrella (i.e., muestairella ) was long regarded as a synonym of Chionodes praeclarella (Herrich-Schäffer, 1854) based on its superficial aspect until its genitalia were eventually examined ( Sauter 1961; Sattler 1961) is telling of how difficult these gelechiid taxa are to recognize from external characters alone. Despite the broad (wide) wings that are characteristic of Agonochaetia , no particular venational features which differed from other Gnorimoschemini were observed. Povolný (2002b) indicated that the venation of members of the tribe did not differ significantly from the common (unspecified groundplan?) pattern exhibited in the Gelechiinae or even in the Gelechiidae , but there is little evidence in most of his works that he examined venation.
About the synonymy of Sautereopsis with Agonochaetia . In the descriptions of Agonochaetia and Sautereopsis, Povolný (1965) did not specify the distinguishing features of both genera. Sattler (1968) synonymized them, arguing that study of their type species did not support their separate generic status and that the discovery of a species with intermediate features, A. intermedia Sattler, 1968 , further supported the generic synonymy. Sattler considered that the difference in the projection of the cucullus (termed ‘dorsal arm of the valva’), which is triangularly produced in A. terrestrella (the type species of Sautereopsis ), simple in A. incredibilis (type species of Agonochaetia ), and intermediate in A. intermedia , is not a feature worthy of generic status. Even though Sattler’s decision implied subjective weighting of character states (differences in shape of a lobe versus absence of lobe), he pointed out other characters uniting the three species then known: gnathos without a hook (= without mesio-distal process); divided valva (=cucullus and sacculus) with the ventral arm (= sacculus) strongly bent; long saccus; paired thin filaments (= glandiductors) arising near the base of aedeagus (= phallus); aedeagus slender with a slightly swollen base. A cucullus with a variously developed ventral lobe is observed in other genera, for example in several Caryocolum and in some Lutilabria , whereas the cucullus is simple in many Gnorimoschemini ( Huemer & Karsholt 2010). Thus it appears that, in order to be meaningful, any character analysis will have to involve a broad array of gnorimoschemine taxa. While we recognize the subjective weighting involved in the current assessment of these characters, we still prefer to retain the generic synonymy pending better and broader character analysis.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Agonochaetia Povolný, 1965
| Landry, Jean-François, Nazari, Vazrick, Bidzilya, Oleksiy, Huemer, Peter & Karsholt, Ole 2017 |
Agonochaetia Povolný, 1965 : 487
| Huemer 2010: 294 |
| Povolny 1965: 487 |
| Povolny 1965: 487 |
Sautereopsis Povolný, 1965 : 488
| Sattler 1968: 119 |
| Povolny 1965: 488 |
| Zeller 1872: 111 |