
Podocotyle pearsei Manter, 1934
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4638.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:E8CE848C-3E8C-45DF-A093-D696D19EBD46 |
|
persistent identifier |
https://treatment.plazi.org/id/51258795-FFB4-8532-D9A1-FE70FD68DB90 |
|
treatment provided by |
Plazi |
|
scientific name |
Podocotyle pearsei Manter, 1934 |
| status |
|
Podocotyle pearsei Manter, 1934 View in CoL
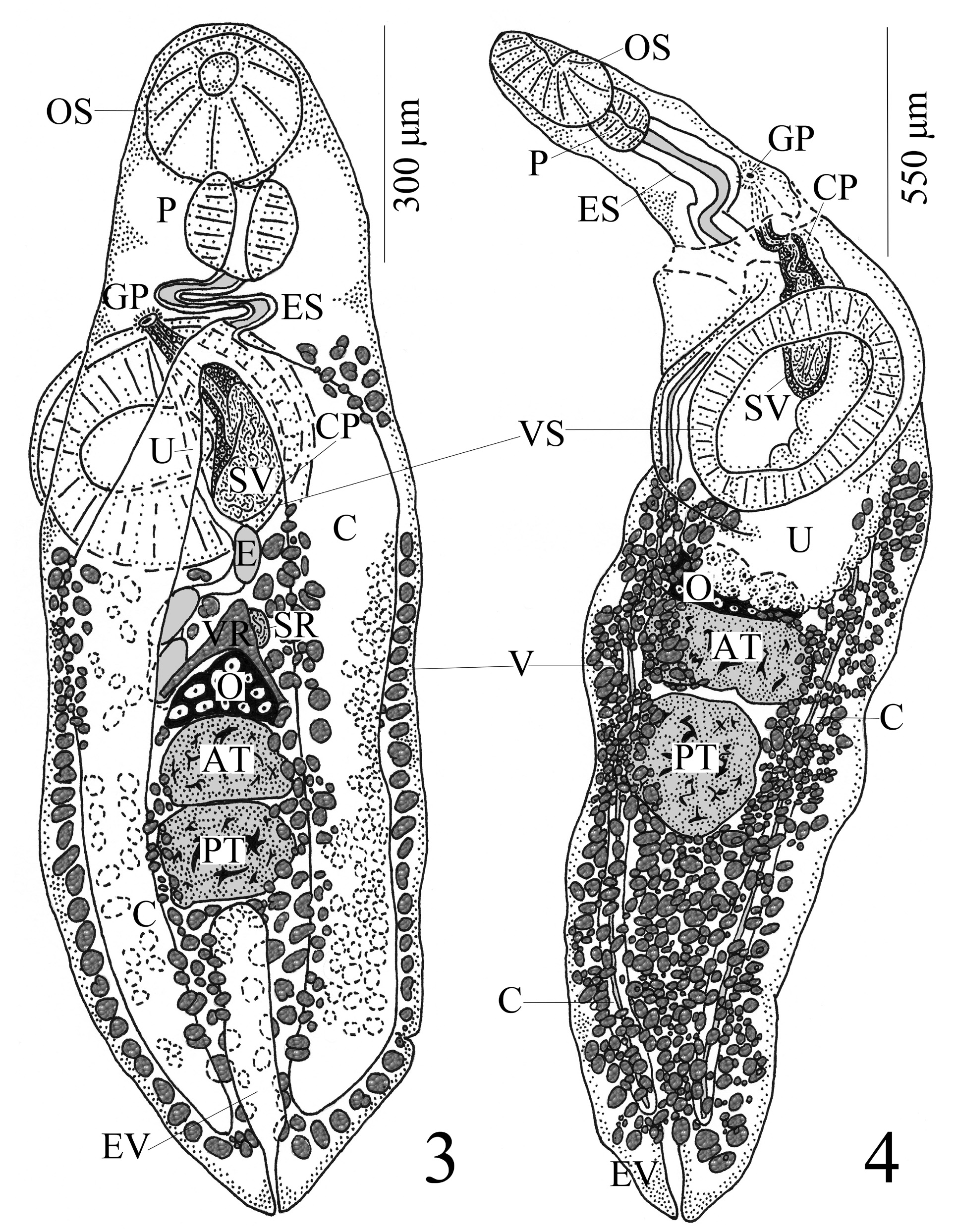
( Fig. 1 View FIGURES 1–2 ; see also Figs 4 View FIGURES 3–4 –6 of Blend et al. 2016)
Synonyms: Podocotyle (Podocotyle) pearsei Manter, 1934 ; Podocotyle sp. n. of Blend (1996) & Blend et al. (2004); Podocotyle sp. of Klimpel et al. (2001, 2009) & Blend et al. (2004).
Host: Vaillant’s grenadier, Bathygadus melanobranchus Vaillant ( Gadiformes : Macrouridae ; Bathygadinae ).
Localities: Western Gulf of Mexico off Tamaulipas, Mexico, 23°32’12” N, 97°03’42” W, depth = 786m, 11/July/1972; Northeastern Gulf of Mexico off Florida, 28°29’58” N, 86°58’09” W, depth = 783–841 m, 13/ May/1985 GoogleMaps .
Site of infection: Intestine.
Prevalence: B. melanobranchus : 4 of 51 (7.8%).
Intensity: B. melanobranchus : 1–38 worms/fish.
Mean intensity: B. melanobranchus : 71/4 = 17.75.
Relative density/abundance: B. melanobranchus : 71/51 = 1.39.
Deposited Specimens: Vouchers (25 individuals; 21 slides) NHMUK 2019.4.12.1 (off Florida) & NHMUK 2019.4.12.2–21 (off Mexico).
Records: 1. Manter (1934, 1940, 1947, 1954b); 2. Park (1937 [key]); 3. Skrjabin & Koval (1958 [key]); 4. Yamaguti (1958, 1971 [listing]); 5. Pritchard (1966 [listing]); 6. Armstrong (1974); 7. Bray (1995 [listing]); 8. Blend (1996); 9. Klimpel et al. (2001, 2009 [listing]); 10. Blend et al. (2004 [listing]); 11. Overstreet et al. (2009 [listing]); 12. Blend et al. 2016; 13. Present study.
Descriptions: 1, 6, 8, 12, 13.
Supplemental descriptive information: [Based on 11 vouchers. Measurements and proportions given in Table 2 View TABLE 2 .] With characteristics of genus. Body elongate oval, widest at junctions of either first and second or second and third 1/3 of body; anterior and posterior ends attenuated to bluntly-rounded extremity, posterior end occasionally tapered to point. Forebody about 1/4–1/3 length of body, attenuate, at times slender. Hindbody either broadens to junction of second and third 1/3 of body or possesses nearly parallel margins before it attenuates to bluntly-rounded or tapered posterior extremity. Tegument aspinose. Pre-oral lobe not observed. Oral sucker subterminal, subspherical to circular to transversely oval, unspecialized. Ventral sucker large, muscular, round to subspherical to oval to transversely oval, unspecialized, sessile to slightly elevated and protuberant, conspicuous border around perimeter, wider than long and almost as wide as entire worm at that level, larger than oral sucker, pre-equatorial at junction of first and second 1/4–1/3 of body. Prepharynx not observed. Pharynx muscular, thick, round to oval (sub-quadrate in 1 specimen), anterior end ventrally-overlapped by oral sucker. Esophagus thick-walled, noticeably sinuous, longer than pharynx. Pharyngeal gland cells proximate to pharynx and anterior esophagus, dark-staining. Intestinal bifurcation either overlapped ventrally by anterior margin of ventral sucker or anterior to ventral sucker by short distance. Ceca thick-walled, parallel or slightly bowed at level of gonads, moderately wide throughout and widest at level of ventral sucker or just posterior to sucker or near junction of second and third 1/3 of body, slightly arcuate far posterior and terminates blindly near posterior extremity.
Testes 2, tandem, median, oval to irregular to oblong in shape, transversely elongate, smooth to slightly indented to lobed, contiguous, intercecal, post-equatorial near junction of second and third 1/3 of body (1 specimen with anterior testis compressed on right side; 1 specimen with circular posterior testis [PT]; 1 specimen with sub-tri- angular-shaped PT; 1 specimen with conspicuous posteriorly-directed lobe on PT). Post-testicular region confined to posterior 1/5–1/4 of body. Cirrus pouch distinct, thin-walled, clavate to “C”-shaped, extends to or near mid-level of ventral sucker along median plane of worm. Seminal vesicle internal, bi-partite; proximal portion overlapped by ventral sucker, large, saccate, occupies majority of proximal portion of cirrus pouch and can be seen to loop back on itself anteriorly-posteriorly before it passes anterior; distal portion thick-walled, narrow and sinuous, runs anteri- orly from proximal portion and anterior to ventral sucker before it turns anterosinistrally – sometimes quite sharply – and terminates. Pars prostatica inconspicuous, thick-walled, tubular, near distal end of cirrus pouch, 28–40 (33) [n = 5] long × 10–24 (17) [n = 5] wide; bleb-like cells large, conspicuous, just inside wall of cirrus pouch between seminal vesicle and distal terminus of male genital system and almost completely fills up lumen of pars prostatica, distributed immediately anterior to and dorsal to anterior margin of ventral sucker. Prostatic gland cells numerous, well-developed, large, dark-stained, distributed throughout cirrus pouch, most numerous at proximal end and in anterodextral portion of cirrus pouch. Genital pore submedian (sinistral), at level of posterior end of lower esophagus (at mid-esophagus level in 1 specimen) and anterior to intestinal bifurcation and ventral sucker, frequently midway between esophagus and left margin of worm but occasionally closer to one or the other. Genital atrium distinct, circular, deep with thick walls.
Ovary 3-lobed, lobes directed either anterior or posterior, transversely elongate, sometimes longitudinally compressed, median to slightly dextral, anterior to and contiguous with anterior testis, in middle third of body. Seminal receptacle canalicular, circular, voluminous, median, dorsally overlaps left, median and/or anterior border of ovary or located directly anterior to ovary. Laurer’s canal present, coiled; distal end opens dorsally, submedian (sinistral) to midline of worm at level of anterior border of ovary and just medial to inner wall of left cecum. Vitelline fields follicular, variable in size, numerous, circular to sub-circular to elongate oval to irregular in shape (rarely subrectangular), circumcecal in uninterrupted lateral bands, extends from posterior extremity anteriorly up to level of posterior margin of ventral sucker (1 specimen with left lateral band of vitelline follicles short of ventral sucker by 96 μm; 1 specimen with right lateral band of follicles that extend anterior to ventral sucker; 1 specimen with cluster of vitelline follicles anterodextral to ventral sucker between esophagus and right margin of worm), encroaches over lateral margins of gonads ventrally and dorsally, not confluent in pre-ovarian region (1 individual with vitellaria confluent here), in space between ovary and anterior testis and in inter-testicular region ( ITR) though follicles can encroach into ITR, confluent in post-testicular region. Vitelline reservoir lemon-shaped to clavate to triangular to sub-triangular in shape, median to submedian (either sinistral or dextral), directly anterior to or dorsally overlaps anterior margin of ovary (1 specimen with vitelline reservoir directly ventral to seminal receptacle). Transverse vitelline ducts run parallel along anterior border of ovary toward lateral margins of worm. Main vitelline duct passes from anterior margin of vitelline reservoir to right to join oviduct which is itself inconspicuous and runs short course anterior to ovary. Oötype immediately anterior to ovary; Mehlis’ gland cells large, anterior to ovary, dorsally overlapped by seminal receptacle, anterior border of Mehlis’ gland cells conspicuous while posterior border of cells more inconspicuous. Uterus either entirely pre-ovarian or extends to mid-level of ovary, widest proximally with many loops and narrows further anteriorly, intercecal and posterior loops pass ventrally over ceca but do not extend lateral to lateral margins of ceca (1 specimen with uterine loops extracecal on left side); uterine loops pro- ceed anteriorly from ovary, pass anterosinistrally over ventral sucker and run alongside and to left of distal end of cirrus pouch to genital pore. Metraterm inconspicuous, thick-walled as it enters genital atrium. Eggs large, collapsed and/or crenulated, operculate, amber, non-filamented, relatively few in number, with nib on one pole; many eggs bloated in proximal uterus (i.e. spherical-shaped); in several specimens eggs present only in proximal uterus with none observed in distal region.
Excretory vesicle I-shaped/tubular, moderately narrow posteriorly and wider anteriorly, extends to posterior testis (1 specimen with excretory vesicle reaching 2/3 distance to posterior testis); excretory pore terminal.
Remarks: This material was assigned to the Podocotylinae within the Opecoelidae based on its possession of the following diagnostic combination of characters: a canalicular seminal receptacle, a well-developed cirrus pouch that is relatively short (extends to or near mid-level of ventral sucker) and encloses an internal seminal vesicle that becomes narrow distally, blind ceca, an intercecal and pre-ovarian uterus, and a deep-sea piscine host ( Cribb 2005; Martin et al. 2018). The present specimens key out to the genus Podocotyle based on its possession of the same diagnostic combination of morphological characters given earlier (see Introduction).
The digenean individuals in this study found in the intestine of B. melanobranchus collected from the western and northeastern Gulf of Mexico compared well to the two earlier published descriptions of P. pearsei : the type description by Manter (1934), who collected P. pearsei from the intestine of the longfin hake, Phycis chesteri Goode & Bean (Phycidae) , from the deep waters of Tortugas, Florida (paratypes - HWML Coll. Access. # 276 & 101928); and a supplemental description of this species by Blend et al. (2016), who documented it from the bullseye grenadier, Bathygadus macrops Goode & Bean, 1885 , the doublethread grenadier, Gadomus arcuatus (Goode & Bean, 1886) and the western softhead grenadier, Malacocephalus occidentalis Goode & Bean, 1885 (Macrouridae) , from deeper waters in the northeastern Gulf of Mexico and the Caribbean Sea off Panama (vouchers - NHMUK 2016.4.28.7–18). While we have identified the material herein as P. pearsei , it should be noted that some minor variation between our material (see our Table 2 View TABLE 2 ) and the type material of Manter (1934) and Blend et al. (2016, Table 3) was observed. On the one hand, body width (655–722 µm [ Manter 1934] and 464–648 µm [ Blend et al. 2016, Table 3] vs 296–480 µm [ Table 2 View TABLE 2 herein] wide at ventral sucker), oral sucker length (218–235 and 224–240 vs 152–216), pharynx length (140–184 vs 92–140), anterior and posterior testis length (128–200 and 184–224 vs 88–120 and 116–180), posterior testis width (280–304 vs 156–264), cirrus pouch length (332–392 vs 226–304), vitelline reservoir length (124–140 vs 50–110) and excretory vesicle length (600–792 vs 320–516) all differed – the earlier-described material being somewhat larger in these features. On the other hand, when compared with the additional specimens (i.e. voucher material) of P. pearsei collected and re-described by Blend et al. (2016, Table 3), the maximum hindbody length (784 vs 1,224) and genital pore distance anterior to the ventral sucker (0–28 vs 44–132) was longer in the present specimens. Forebody length as a percentage of body length was higher in the present material, too (24.8%–34.3% [our Table 2 View TABLE 2 ] vs “less than 1/4 body length” [ Manter 1934, p. 289] and 21.1%–26.0% and 15.2%–27.9% [ Blend et al. 2016, Table 3]). We noted that the excretory vesicle of P. pearsei was reported earlier to extend to the ovary; however, in the current material it extends only to the posterior testis. This might be due to the difficulty of observing the anterior extent of the excretory vesicle in this material as it was often concealed by the testes and vitelline fields (i.e. excretory length measured in only 7 specimens – see Table 2 View TABLE 2 ). While the present specimens of P. pearsei were cold-fixed (see Table 2 View TABLE 2 ), all remaining absolute measurements of features as well as conventional allometric values and ratios were similar to those of conspecific, heat-fixed specimens published earlier including egg size (80–100 × 42–62 [our Table 2 View TABLE 2 ] vs 96–105 × 39–45 [ Manter 1934, p. 290] and 84–104 × 38–52 and 90–104 × 40–64, respectively [ Blend et al. 2016, Table 3]); however, eggs in the present specimens were fewer in number and had a nib on one end – this latter feature was not noted in earlier descriptions of P. pearsei . In addition, Manter (1934) mentioned that the posterior testis of P. pearsei was “more or less triangular”; indeed, one of our specimens herein had a sub-triangular-shaped posterior testis. The distinctive loop of the seminal vesicle in the current specimens of P. pearsei was noted as well (see Manter 1940, p. 386; Blend et al. 2016, p. 505).
It is important to note that among the current specimens of P. pearsei included in Table 2 View TABLE 2 , some intra-specific variation in morphological features was observed. One specimen had a sub-quadrate-shaped instead of round to oval pharynx. Most specimens possessed testes that were oval to irregular to oblong in shape, transversely elongate, and smooth to slightly indented to lobed in texture; however, one specimen had an anterior testis that was compressed on the right side, two specimens possessed either a sub-triangular or circular-shaped posterior testis and a fourth specimen contained a conspicuous, posteriorly-directed lobe on the posterior testis. The genital pore was observed at the level of the posterior end of the lower esophagus, but in one specimen, it was at mid-esophagus level (see Blend et al. 2016, p. 505 where variability in genital pore position relative to the esophagus and intestinal bifurcation in P. pearsei also was noted). We noted the vitelline follicles rarely to have a sub-rectangular shape as opposed to a circular to sub-circular to elongate oval to irregular shape, and while the anterior extent of the vitelline fields was to the level of the posterior margin of the ventral sucker in the majority of specimens observed, we noted three individuals with variability in this feature. The left lateral band of vitelline follicles fell short of the ventral sucker by 96 μm in one specimen, the right lateral band of follicles extended anterior to the ventral sucker in a second specimen, and there was a cluster of vitelline follicles anterodextral to the ventral sucker between the esophagus and right body margin in another worm. One specimen had confluent vitelline fields in the pre-ovarian region and another possessed a vitelline reservoir directly ventral to the seminal receptacle and not directly anterior to or dorsally overlapping the anterior margin of the ovary. While the uterus of P. pearsei is intercecal, one of our specimens possessed a uterus with extracecal loops on its left side.
This is the third published report containing descriptive information of P. pearsei from undocumented / un-deposited materials (see Blend et al. 2016, p. 508).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Podocotylinae |
|
Genus |