
Astiotrema impletum ( Looss, 1899 ) Looss, 1900
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4991.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:585A1043-224B-488B-ADBA-47DE8DE27869 |
|
DOI |
https://doi.org/10.5281/zenodo.5088897 |
|
persistent identifier |
https://treatment.plazi.org/id/5046879B-FFD3-A50D-4D86-FC5A5CD2FB9D |
|
treatment provided by |
Plazi |
|
scientific name |
Astiotrema impletum ( Looss, 1899 ) Looss, 1900 |
| status |
|
Astiotrema impletum ( Looss, 1899) Looss, 1900
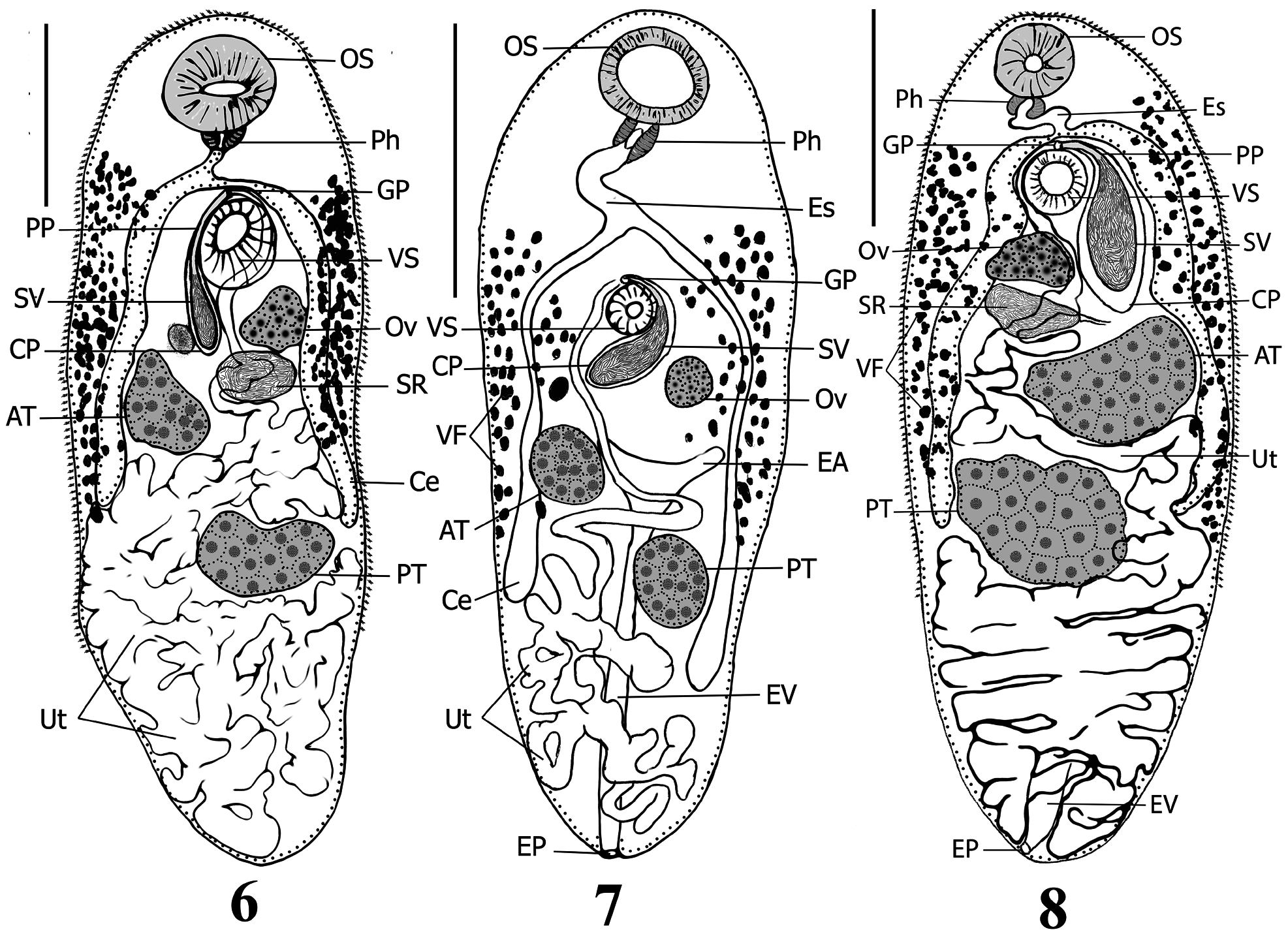
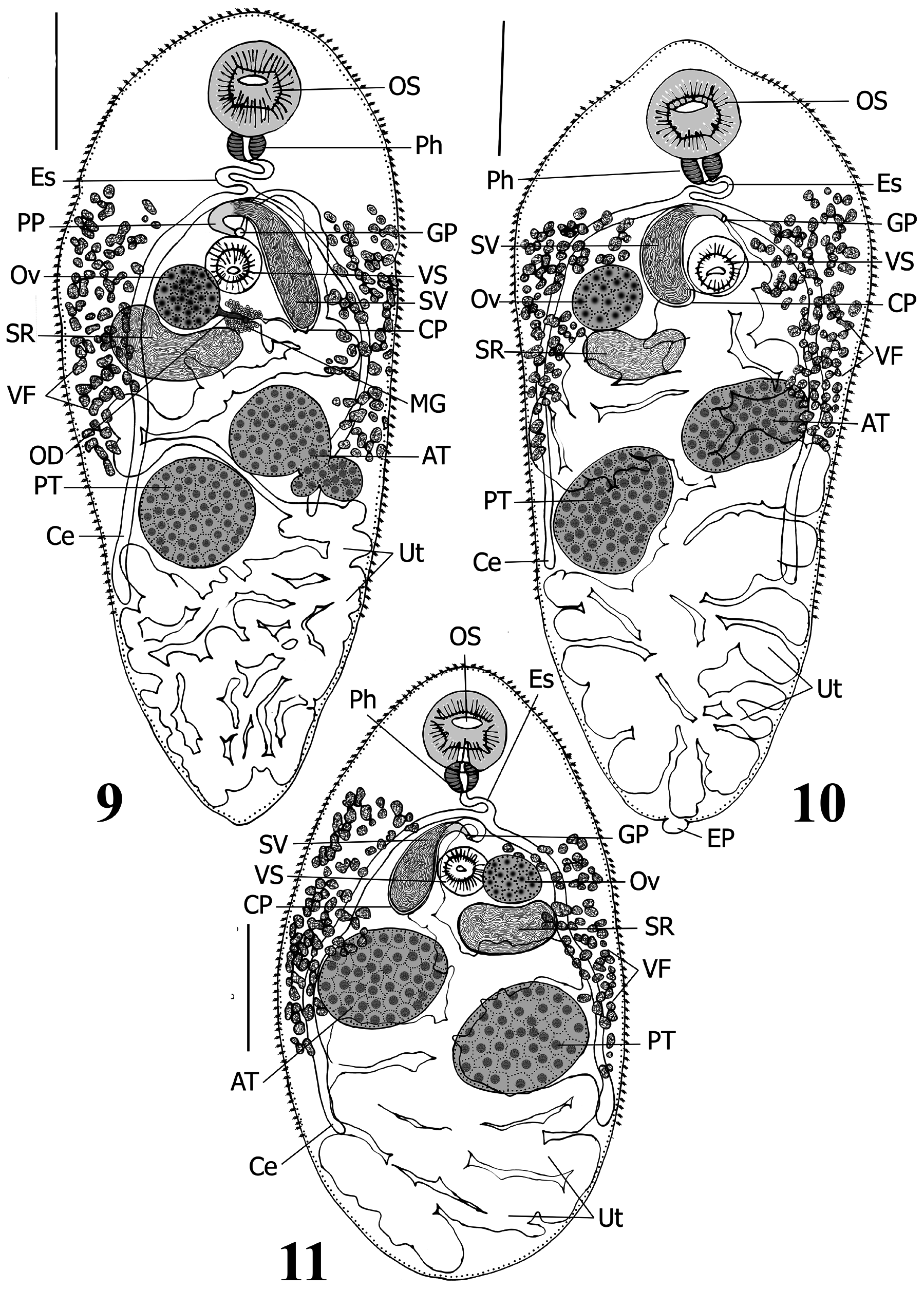
( Figs. 6–11 View FIGURES 6–8 View FIGURES 9–11 )
(Syns.: Astia impleta Looss, 1899 ; Tremiorchis varanum Verma, 1930 n. syn.; Astiotrema sudarshani Mukherjee & Ghosh, 1970 n. syn.; Astiotrema varanum [Verma, 1930] Mukherjee & Ghosh, 1970 n. syn.; Astiotrema varanusi Gupta & Jahan, 1978 n. syn.)
Host: Tetraodon lineatus Linnaeus ; Tetraodontiformes : Tetraodontidae ; Globe fish.
Locality: From the River Nile at Qena, Egypt, 26°08’N, 32°42’E –26°09’N, 32°41’E; depth = 0.60–7.50 m; June/2018 & July/2018 GoogleMaps .
Site of infection: Intestine.
Prevalence: 3 of 8 Tetraodon lineatus (37.5% infected).
Intensity: 20–45 worms/host specimen.
Mean intensity: 31.7 (95/3) worms.
Relative density/abundance: 11.9 (95/8) worms.
Deposited specimens: BM(NH) vouchers NHMUK.2021.5.26.1-3; SVU vouchers (SVU) 21.18.Ai01–Ai05.
Records (see Table 1 View TABLE 1 ): 1. Looss (1899); 2. Odhner (1911); 3. Verma (1930); 4. Yeh & Fotedar (1958); 5. Fischthal & Kuntz (1963); 6. Mukherjee & Ghosh (1970); 7. Gupta & Jahan (1978); 8. El-Naffer et al. (1984); 9. Hamada (2002); 10. Present study.
Supplemental description. [Based on 17 ovigerous specimens] Body elongate-oval with almost parallel margins, range 2,211 –3,578 (mean 2,860) × 1,116 –1,540 (1,299); maximum width at area anterior to anterior testis to anterior limit of ventral sucker level, 38.4%–57.8% (45.8%) of body length. Anterior end broadly rounded; posterior end less broadly rounded; pre-oral lobe distinct, short, 82–120 (105) long, 2.8%–4.6% (3.7%) of body length. Forebody 617–881 (773) long, 20.8%–33.8% (27.4%) of body length, gradually narrows near anterior end. Hindbody wider and longer than forebody, gradually narrows in posterior third of body. Tegument armed with numerous small, scale-like spines, spines 8–12 µm long; concentrated in forebody, sparser gradually towards posterior limit of level of posterior testis, post-testicular area devoid of spines. Oral sucker globular, sub-terminal, unspecialized with conspicuous large central mouth opening, 290–412 (346) × 298–410 (356), 10.9%–13.3% (12.2%) of body length. Ventral sucker sessile, unspecialized, globular, pre-equatorial, distinctly smaller than oral sucker, 168–277 (211) × 171–286 (222), 6.9%–8.3% (7.4%) of body length. Suckers well-separated by a long distance 230–405 (343) long, 7.0%–17.3% (12.3%) of body length. Oral sucker/ventral sucker length and width ratios 1:0.55–0.67 (1:0.61) & 1:0.57–0.70 (1:0.62), respectively. Prepharynx absent. Pharynx muscular, transversely oval, well-developed, narrower and smaller than both suckers, not papillate, 96–136 (115) × 95–163 (144), 3.4%–5.0% (4.1%) of body length. Pharynx/oral sucker length and width ratios 1:2.51–3.35 (1:3.02) & 1:2.22–3.71 (1:2.50). Pharynx/ventral sucker length and width ratios 1:1.60–2.25 (1:1.85) & 1:1.29–2.38 (1:1.56). Prebifurcal distance 568–725 (653) long, 19.8%–29.3% (23.1%) of body length. Esophagus distinct, thin, long, sigmoid [n=15], 144–398 (252) long, 4.9%–12.8% (8.9%) of body length, rarely contracted [n=2]. Intestinal bifurcation in mid-forebody, at distance 568–725 (653) from anterior extremity and 19.8%–29.3% (23.1%) of body length, separated from ventral sucker by small distance 20–195 (106), 0.6%–6.1% (3.8%) of body length. Ceca 2, simple, blind, arcuate, subequal in length, equal in width, pass posteriorly along lateral margins of worm to end at level of posterior testis or slightly further, at distance 414–1,041(771) from posterior extremity, represents 18.7%–31.6% (26.6%) of body length.
Testes 2, consistently globular to oval, equally-sized, entire, oblique [n=16], rarely opposite [n=1], occupy middle third of body, widely separated from posterior end, separated by narrow inter-testicular space, 19–164 (83) long, 0.7%–6.8% (2.9%) of body length; anterior testis 313–558 (428) × 357–552 (432), 12.8%–17.5% (14.9%) of body length; posterior testis 396–618 (483) × 338–560 (444), 13.4%–19.4% (17.0%) of body length. Pre-testicular distance 1,020 –1,568 (1,295) long, 42.2%–53.7% (45.5%) of body length. Post-testicular space approximately represents last quarter of body, 475–1,134 (825) long, 21.5%–34.2% (28.5%) of body length. Cirrus-pouch large, claviform, curved to C-shaped, 382–751 (572) × 108–217 (165), 14.8%–25.3% (19.9%) of body length, extends posteriorly around left edge of ventral sucker [n=13] or occasionally around right edge [n=4] to ovarian level; encloses well-developed prostatic cells, ejaculatory duct, pars prostatica and seminal vesicle. Internal seminal vesicle broadly saccular, unipartite, occupies about two thirds of cirrus-pouch, 215–508 (375) × 108–217 (165), 9.7%– 18.9% (13.1%) of body length, connects to well-developed, short, thin-walled, straight, pars prostatica. Ejaculatory duct short, indistinct from pars prostatica, extends slightly anterior to ventral sucker before it turns posteriorly to enter shallow genital atrium. Genital pore post-bifurcal in forebody, median [n=12] to slightly sinistro-submedian [n=5], mid-way between ventral sucker and intestinal bifurcation [n=10] to immediately anterior to ventral sucker [n=3] or immediately ventral to intestinal bifurcation [n=4], 643–830 (726) from anterior end or 21.3%–32.4% (25.7%) of body length.
Ovary entire, rounded, 165–258 (232) × 167–286 (246), 7.0%–9.2% (8.2%) of body length, pre-testicular, distinctly separated from anterior testis, dextral to ventral sucker and either separated by a slight distance or contiguous to it [n=13]; in some specimens, ovary separated by a short transverse distance 81–218 (156) long, occupied by cirrus-pouch [n=4]. Preovarian region 748–1,087 (924) long, 28.9%–41.4% (32.6%) of body length from anterior end. Mean testes/ovary length and width ratios 1:0.40–0.61 (1:0.51) & 1:0.45–0.67 (1:0.57), respectively. Seminal receptacle conspicuous, broadly saccate, 155–508 (363) × 134–250 (190), situated in area posterior to ovary and may occupy space between ovary and anterior testis. Öotype median, surrounded by a large mass of Mehlis’ gland cells; both in small area proximate to posterior margin of ventral sucker, anterior margin of anterior testis and either sinistral or dextral margins of seminal receptacle and cirrus-pouch. Uterus well-developed, highly coiled, intercecal, occupies entire post-testicular area, extends anteriorly and fills inter-testicular space to level of posterior margin of ventral sucker. Metraterm thin-walled, extends along right side of ventral sucker, reaches slightly anterior to genital pore level then turns posteriorly to enter genital atrium. Vitellarium mainly extracaecal; follicles distributed dorsal and ventral to ceca, relatively numerous, moderate in size, subglobular to oval; fields extend along lateral margins from near level of pharynx or slightly posterior to it and anterior to intestinal bifurcation to terminate near posterior testis level; pre-vitelline region 447–728 (607) long, 17.4%–27.1% (21.4%) of body length; post-vitelline region 699–1,671 (1,136) long, 31.6%–46.7% (39.2%) of body length. Small vitelline ductulus unite at anterior extremity of each vitelline field forming one transverse vitelline duct that extends from each side then unites in a small median vitelline reservoir directly anterior to ventral sucker or overlaps it dorsally. Eggs numerous, oval, 38–46 (42) × 13–20 (16) [n=21], operculate, thin-shelled, without filaments.
Excretory vesicle Y-shaped, stem extends anteriorly, passes between testes and bifurcates posterior to ovary into two short arms; excretory pore terminal.
Remarks: Comparisons among records of A. impletum from the type host, the globe fish, Tetraodon lineatus (Syn. Tetraodon fahaka Hasselquist ) (see Looss 1899; Odhner 1911; Yeh & Fotedar 1958; Fischthal & Kuntz 1963; El-Naffar et al. 1984), and the present material reveal some morphological variation within this species: (i) genital pore position relative to the intestinal bifurcation differentiated slightly from pre-bifurcal ( Looss 1899) to bifurcal ( Yeh & Fotedar 1958; Present study) to post-bifurcal ( Odhner 1911; Fischthal & Kuntz 1963; El-Naffar et al. 1984; Present study); (ii) genital pore position varied from median ( Looss 1899; Fischthal & Kuntz 1963; El-Naffar et al. 1984; Present study) to slightly submedian and sinistral ( Odhner 1911; Present study); (iii) testes either well-separated ( Looss 1899; Odhner 1911; Yeh & Fotedar 1958; El-Naffar et al. 1984; Present study) or contiguous ( Fischthal & Kuntz 1963; Present study); (iv) testes either oblique ( Looss 1899; Odhner 1911; Yeh & Fotedar 1958; Fischthal & Kuntz 1963; El-Naffar et al. 1984; Present study) or opposite ( El-Naffar et al. 1984; Present study); (v) testes lobulation and shape range from occasionally slightly lobed and almost symmetrical ( Odhner 1911) to entire and more diagonal in position ( Looss 1899; Yeh & Fotedar 1958; Fischthal & Kuntz 1963; El-Naffar et al. 1984; Present study); and (vi) the position of the cirrus-pouch relative to the ventral sucker and ovary varies from either dextral to both ventral sucker and ovary (i.e., ventral sucker situated between ovary and cirrus-pouch) (see Looss 1899; Odhner 1911; Yeh & Fotedar 1958; El-Naffar et al. 1984; Present study) or confined between right edge of ovary and left edge of ventral sucker (i.e., ventral sucker sinistral to both cirrus-pouch and ovary) (see El-Naffar et al. 1984; Present study). The pre-bifurcal position of the genital pore in A. impletum as stated by Looss (1899) is inconsistent with all reported subsequent records from the type host. We suspect that Looss (1899) was mistaken and that the true position of the genital pore is either post-bifurcal or occasionally bifurcal. We interpret the remaining morphological variation listed above, in part, to intra-specific variation and/or differences in parasite preparation and the potential effect of change in the host locality. Characteristics of A. impletum that we consider consistent are as follows: (i) oral sucker distinctly larger than ventral sucker, (ii) vitelline fields extend from pharyngeal level to the anterior extent of posterior testes level, (iii) ceca terminate at posterior limit of posterior testes level, (iv) ovary either contiguous to ventral sucker or posterior to it by a very small distance, and (v) a sigmoid esophagus.
Verma (1930) reported T. varanum from the small intestine of two reptilians, the Bengal or Indian monitor, Varanus bengalensis (Daudin) , and the desert monitor Varanus griseus (Daudin) ( Squamata : Varanidae ), from Allahabad, India. Mukherjee & Ghosh (1970) noted that T. varanum resembles species of Astiotrema more than Tremiorchis , particularly in regard to the posterior extent of the intestinal ceca, the position of the testes and the sigmoid nature of the excretory vesicle. Consequently, they transferred T. varanum to Astiotrema as A. varanum . Astiotrema varanusi represents another closely related species collected from the intestine of an unidentified species of Varanus Merrem from Lucknow, India ( Gupta & Jahan 1978). The two species have identical morphology as well as sharing the same locality ( India) and host group (monitors), thus we synonymize A. varanusi with A. varanum . Furthermore, A. varanum and its synonym A. varanusi are morphologically identical with A. impletum ; we could not find any distinct characteristics differentiating them except A. impletum was reported from a fish host (globe fish) and A. varanum is known from monitor lizards. The fresh and brackish water globe fish is widely spread throughout Africa, particularly in large rivers, open water, weed beds and vegetated fringes ( Froese & Pauly 2021); whereas, monitor lizards are native to Africa, Asia, and Oceania ( Uetz et al. 2021). Both the globe fish and monitor lizards are entirely carnivorous; the globe fish feeds on mollusks ( Froese & Pauly 2021) and monitor lizards consume a wide variety of food such as insects, crustaceans, arachnids, myriapods, mollusks, fish, amphibians, reptiles, birds, and mammals ( Pianka et al. 2004). As A. varanum and A. impletum share the same morphology as well as host geographical distribution (Africa) and feeding habitats (consume mollusks), we conclude A. varanum as another synonym of A. impletum .
Mukherjee & Ghosh (1970) described A. sudarshani for specimens collected from the intestine of the Southeast Asian or Asian common toad, Duttaphrynus melanostictus (Schneider) (Syn. Bufo melanostictus Schneider ) ( Anura : Bufonidae ), from Birati, West Bengal, India. We note no distinct characteristics differentiating A. sudarshani from A. impletum except that it was recorded from anurans. In addition, D. melanostictus is a completely carnivorous toad that occurs widely in Asia; it consumes a wide variety of food, mainly arthropods and mollusks ( Berry & Bullock 1962). Given that A. sudarshani and A. impletum have indistinguishable morphology, their hosts share feeding habitats (consume mollusks) and A. impletum (Syn. A. varanum ) was reported earlier from the same geographical area (Asia), we synonymize A. sudarshani with A. impletum .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |