
Choeroniscus minor (Peters)
|
publication ID |
https://doi.org/ 10.5281/zenodo.4545052 |
|
DOI |
https://doi.org/10.5281/zenodo.4550899 |
|
persistent identifier |
https://treatment.plazi.org/id/4F19FC10-FFA3-FF94-FD0E-218CFC608F08 |
|
treatment provided by |
Plazi |
|
scientific name |
Choeroniscus minor (Peters) |
| status |
|
Choeroniscus minor (Peters) View in CoL
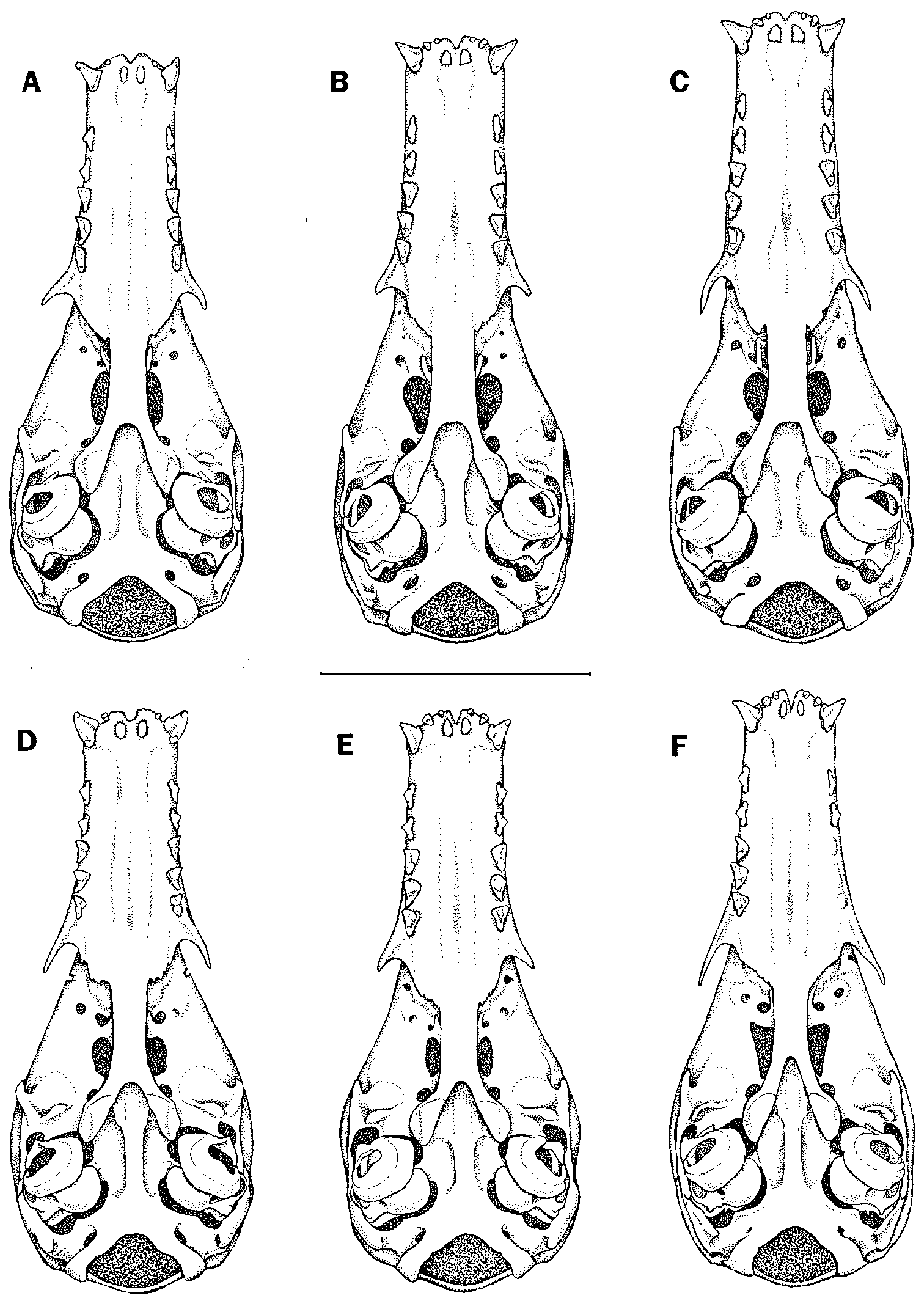
Figure 24 View Fig
VOUCHER MATERIAL: 8 females (AMNH *266120, *266121, *266123, *266377, *267153; MNHN *1998.668, 1998.669, *1998.670) and 5 males (AMNH *266124, *267946, *267947, *267948; MNHN 1998.671); see tables 13 and 14 for measurements.
IDENTIFICATION: The complex taxonomic history of Choeroniscus , particularly of the smaller species originally placed in the genus Choeronycteris , is unfortunately relevant for understanding the correct identification of our Paracou vouchers.
Choeronycteris minor was first described by Peters (1868) based on a specimen from Surinam. Although Goodwin and Greenhall (1961) stated that the type was destroyed in World War II, Husson (1962) reported that Peter’s original specimen was, in fact, still intact at the Stuttgart museum. Husson (1962) published measurements and a photograph of the holotype (SMN 441), and also provided a detailed description. A second small species, Choeronycteris intermedia , was described by Allen and Chapman (1893) based on specimens from Trinidad, and Tho mas (1912a) named a third, Choeronycteris inca , based on a specimen from Peru. Thomas (1928a) subsequently subdivided Choeronycteris , placing minor , intermedia , and inca in his new genus Choeroniscus .
6 We follow Handley’s (1984) spelling of the specific epithet, which Koopman (1993) and most other authors have incorrectly spelled ‘‘ caudifer.’’
Considerable confusion has surrounded the use of these three specific epithets (Thomas, 1912a; Sanborn, 1954; Goodwin and Greenhall, 1961; Husson, 1962; Koopman, 1978; and Williams and Genoways, 1980a), much of it the result of small available sample sizes and marked sexual dimorphism ( Genoways et al., 1973; Koopman, 1978). The latter factor is crucial because the holotypes of minor and inca are males, while that of intermedius is a female. Conflicting reports concerning the morphology and sex of the supposedly lost holotype of minor only served to compound the real biological problem, whether these names really represent three or fewer species.
There is general agreement that Allen and Chapman (1893) named their Tinidadian material as a new species in part because several measurements of minor were incorrectly reported by Peters (1868). Thomas (1912a: 404) observed that
The Trinidad bat described by Allen and Chapman as Ch. intermedia is very probably the same as Ch. minor , as those authors were deceived by Peter’s impossible measurement of 11 mm. for the calcar, an organ drawn as about 5 mm. long in the more recently published plate of the latter species...
Thomas (1912a) then described inca based on dental proportions (length and width of the premolars and molars) in which it supposedly differed from minor . He also noted that the skull of inca was larger and broader than that of minor , but this comparison was apparently based on a specimen of minor other than the holotype, as measurements of the latter do not confirm his observation (table 13).
Most recent authors have followed Koopman (1978) in regarding inca as a junior synonym of minor , and intermedius as a potentially distinct species. Koopman’s (1978: 8) conclusion that two species could be recognized was based on observations concerning dental variation and length of the rostrum:
Study of a number of skulls of Choeroniscus from Trinidad, British Guiana, Brazil, Peru, and Ecuador [has] convinced me that two species can be distin guished... by rostral length, which is reflected in maxillary tooth row length. Thomas (1912[a]) distinguished inca from minor solely by the relative sizes of the premolars and molars. On the basis of these skulls, I can see considerable variation in this character, but no clearcut distinction between two forms. The species with the longer rostrum would then be called C. minor ( inca a synonym) and the species with the shorter rostrum would stand as C. intermedia [sic].
Koopman (1994) gave the range of minor as extending from coastal Ecuador to the mouth of the Amazon, northward to eastern Venezuela, and southward to northwestern Bolivia. Although most specimens of small Choeroniscus recently collected in the Guianas have been identified as intermedius (e.g., by Brosset and CharlesDominique, 1990), a longmuzzled female from Guyana (AMNH 140471) was referred to minor by Williams and Genoways (1980a).
In attempting to identify our material from Paracou, we examined the holotype of intermedius ; Husson’s (1962) measurements, photographs, and description of the holotype of minor ; Thomas’s (1912a) description of the holotype of inca ; specimens referred to intermedius and minor in the AMNH; and measurements of additional specimens referred to intermedius and minor by Genoways et al. (1973), Genoways and Williams (1980a), Ochoa and Fernández (1982), Brosset and CharlesDominique (1990), and Anderson (1997). Although we kept males and females separate, and investigated variation in size, skull shape, and dental morphology, we found no consistent differences among named forms of small Choeroniscus . For example, dental morphology in our series of 13 specimens from Paracou is highly variable, with some individuals having much shorter or longer teeth than others, others having narrower or broader teeth, etc. These differences form no consistent pattern and do not appear to be correlated with body size or with rostral proportions. Because our sample includes individuals with both ‘‘ minor type’’ and ‘‘ inca type’’ teeth (sensu Thomas, 1912a), as well as others with intermediate conditions, we follow Koopman (1978, 1993, 1994) in concluding that minor and inca are conspecific.
Dental morphology has never been used to separate minor (or inca ) from intermedius . Instead, measurements of the skull, rostrum, and maxillary toothrow appear to be the only characters so employed in recent years ( Koopman, 1978; Williams and Genoways, 1980a). Rather than confirming that minor and intermedius are distinct species, our comparisons of measurement data show the opposite. Whereas measurements of our female specimens from Paracou broadly overlap those of the female holotype of intermedius , measurements of our males overlap those of the male holotypes of minor and inca (tables 13, 14). For both sexes, mea surements of our sample correspond closely to those reported for a large series (10 males and 26 females) of topotypical intermedius from Trinidad ( Genoways et al., 1973). Measurements of other specimens from Venezuela and the Guianas (reported by Genoways and Williams [1980a], Ochoa and Fernández [1982], and Brosset and CharlesDominique [1990]) and Bolivia (Anderson, 1997) further fill the morphometric gap that Koopman (1978) observed between his small series of minor and intermedius . Considering the possiblity that our largest voucher specimens
might represent minor and the smallest intermedius , we carefully examined variation in external and craniodental morphology within our series, but ultimately found no evidence that the Paracou material represents more than one species. In our view, specimens identified as intermedius in the literature simply represent the lower end of a normal range of withinspecies size variation, while those identified as minor and inca represent the upper end of this range. We therefore conclude that these epithets represent a single species for which the oldest available name is Choeroniscus minor (Peters, 1868) . Recognition of subspecies appears to be unjustified based on the data currently at hand.
FIELD OBSERVATIONS: We captured 13 individuals of Choeroniscus minor at Paracou, of which 7 were taken in groundlevel mist nets and 6 at roosts. Three individuals were netted in swampy primary forest, three in creekside primary forest, and one in a manmade clearing. One roosting group of six bats was found on the dark underside of a fallen tree with large buttresses (similar to that shown in fig. 17) in welldrained primary forest; of this group, one adult male and four adult females were captured, and one bat escaped. Another roosting bat, a solitary adult male, was found hanging beneath the undercut bank of a dry streambed in welldrained primary forest.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Glossophaginae |
|
Genus |