
Obama nungara
|
publication ID |
https://doi.org/ 10.1111/zoj.12358 |
|
DOI |
https://doi.org/10.5281/zenodo.5621678 |
|
persistent identifier |
https://treatment.plazi.org/id/4E2E87C6-2126-B63A-AA4C-FEC6CC7A173C |
|
treatment provided by |
Plazi |
|
scientific name |
Obama nungara |
| status |
|
OBAMA NUNGARA View in CoL SP. NOV.
SYNONYMY
Obama sp. 6 (in Carbayo et al., 2013) View Cited Treatment .
Obama marmorata (in Lago-Barcia et al., 2015) View in CoL .
Etymology: The specific epithet refers to the Tupi (indigenous Brazilian tribe) name nungara , meaning similar, alike ( Tibiriçá, 1984). It refers to the resemblance of the species to Obama marmorata .
Type locality: Paulo Lopes, Santa Catarina, Brazil.
Diagnosis: Species of Obama up to 70 mm long, 5 mm wide; brown dorsum covered with small longitudinal black striae. Some specimens may be almost black. Pale midline may be present. Eyes occupying a lateral band on each side of the body 1/3rd of body width. Relative mouth/body length c. 60%. Relative gonopore/ body length c. 77%. Copulatory apparatus 3.26 ± 0.75 mm from anterior to posterior. Dorsal insertion of the penis anterior to the level of the gonopore. Penis orientated backwards to the left side of the body. Necks of glands producing pinkish granules (may also be blueish) joined in conspicuous bundles in the penis. Male atrium provided with a sheath-like large fold on the right side, separating the male from the female atrium. Ring-shaped surface of male atrium crossed by cyanophilic glands extending from the anteroventral to the postero-dorsal side of the male atrium. Lining epithelium of female atrium not stratified in appearance.
Molecular diagnosis: this species includes all populations that cluster with individuals MZUSP PL 1066, MZUSP PL 1067, MZUSP PL 1598, MZUSP PL 1599, Harvard- IZ 59101, NHMUK 2008.4.1.3 and NHMUK 2008.4.1.4 ( Table 1) from this study, with significant support in an adequate molecular delimitation model.
Type material
Holotype MZUSP PL 1594 (field number F3190): Parque Estadual Serra do Tabuleiro, Paulo Lopes, Santa Catarina, Brazil (27°57′35.6″S, 48°45′27.4″W). F. Carbayo et al. coll. 9/Jan/2009. A portion 7.5 mm long including pharynx, sagittal sections on 24 slides; a portion 10 mm long including copulatory apparatus, sagittal sections on 19 slides. Paratype MZUSP PL 1066 (field number F3191): Parque Estadual Serra do Tabuleiro, Paulo Lopes, Santa Catarina, Brazil (27°57′35.6″S, 48°45′27.4″W). F. Carbayo et al. coll., 9/Jan/2009. Anterior end 5 mm long, transverse sections at 7 μm on 16 slides; a portion 9 mm long behind anterior end, containing ovaries, horizontal sections at 7 μm on 11 slides; a portion 8 mm long, horizontal sections at 7 μm on 14 slides; the pre-pharyngeal region, 3 mm long, transverse sections at 7 μm on ten slides; a portion 6 mm long including pharynx, sagittal sections at 7 μm on 53 slides; a portion 9 mm including the copulatory apparatus, sagittal sections at 7 μm on 30 slides; a 5-mmlong tail kept frozen in absolute ethanol. Paratype MZUSP PL 1595 (field number F3227): Parque Estadual Serra do Tabuleiro, Paulo Lopes, Santa Catarina, Brazil (27°57′41″S, 48°45′30″W). F. Carbayo et al. coll., 9/Jan/2009. A portion 8 mm long including the pharynx, sagittal sections on 11 slides; a portion 8 mm long including the copulatory apparatus, sagittal sections on 14 slides. Paratype MZUSP PL 1067 (field number F3247): Parque Estadual Serra do Tabuleiro, Paulo Lopes, Santa Catarina, Brazil (27°58′53″S, 48°44′53″W), F. Carbayo et al. coll., 13/ Jan/2009. Anterior end 6 mm long, transverse sections at 7 μm on nine slides; a portion 10 mm long behind anterior end, containing ovaries, horizontal sections at 7 μm on 19 slides; a portion behind, 5 mm long kept frozen in absolute ethanol; pre-pharyngeal region 4 mm long, transverse sections on 13 slides; a portion 9 mm long including pharynx, sagittal sections at 7 μm on 23 slides; a portion 8 mm long including the copulatory apparatus, sagittal sections at 7 μm on 21 slides; a 7-mm-long end portion preserved in 80% ethanol. Paratype NHMUK 2013.9.5.1: Torroella de Fluvià, Girona, Spain (42°10′29.4″N, 3°02′41.0″E) M. Riutort, coll. 2010. A portion anterior to the pharynx 5 mm long, transverse sections at 10 μm on six slides; a portion 10 mm long including copulatory apparatus, sagittal sections at 15 μm on 12 slides. Remainder preserved in absolute alcohol. Paratype MZUSP PL 1596 (field number F4649): Torroella de Fluvià, Girona, Spain (42°10′29.4″N, 3°02′41.0″E). M. Riutort, coll., 1/Sep/ 2010. A portion 8 mm including the copulatory apparatus, sagittal sections at 7 μm on 19 slides. Paratype MZUSP PL 1597 (field number F4650): Torroella de Fluvià, Girona, Spain (42°10′29.4″N, 3°02′41.0″E). M. Riutort, coll., 1/Sep/2010. The anterior end 5 mm long, transverse sections at 7 μm on 13 slides; a portion 21 mm including pharynx and copulatory apparatus, sagittal sections at 7 μm on 64 slides. Paratype MZUSP PL 1599 (field number F6102): Torroella de Fluvià, Girona, Spain (42°10′29.4″N, 3°02′41.0″E). Riutort & Carbayo, coll., 2/Mar/2014. A portion 8 mm long including pharynx, sagittal section at 7 μm on 21 slides; a portion 8 mm long including copulatory apparatus, sagittal sections at 7 μm on 14 slides. Paratype MZUSP PL 1598 (field number F6098): Torroella de Fluvià, Girona, Spain (42°10′29.4″N, 3°02′41.0″E). M. Riutort, coll., 19/Jan/2014. The anterior end 15 mm long, horizontal sections at 7–14 μm on 19 slides; the prepharyngeal region, 2 mm long transverse sections at 10 μm on five slides; a portion 7 mm long including pharynx, sagittal sections at 10 μm on nine slides; a portion 7 mm including the copulatory apparatus, sagittal sections at 7–12 μm on 11 slides; a ∼ 7 mm long piece of the posterior tip kept frozen in absolute ethanol. Paratype NHMUK 2008.4.1.1: Guernsey, UK (49°27′21″N, 2°34′16″W). T. Brokenshire & N. Clark, coll., 17/Mar/2008. Anterior end approximately 10 mm long, sagittal sections at 15 μm on 15 slides engraved G1 1–15; a portion behind the anterior end about 5 mm long, transverse sections at 10 μm on 13 slides engraved G2 1–13; a portion about 10 mm long containing the pharynx, sagittal sections at 10 μm on 25 slides engraved G3 1–25; the posterior portion about 15 mm long including the copulatory apparatus, sagittal sections at 15 μm on 16 slides engraved G4 1–16. Paratypes NHMUK 2008.4.1.2, NHMUK 2008.4.1.3 and NHMUK 2008.4.1.4: Guernsey, UK (49°27′21″N, 2°34′16″W). T. Brokenshire & N. Clark, coll., 17/Mar/ 2008., preserved in absolute ethanol. Paratype NHMUK 2010.10.29.1: Fordham, Cambridgeshire, UK (52°18′31″N, 0°22′57″E). L. Parker & R. Gordon, coll., July 2009. Preserved in absolute ethanol. Small posterior portion removed for molecular analysis.
Additional material studied
Specimens F6101, F6105, F6106 and F6100: Torroella de Fluvià, Girona, Spain (42°10′29.4″N, 3°02′41.0″E). M. Riutort and F. Carbayo, coll., 19/Mar/2014. NHMUK 2013.9.5.2: Torroella de Fluvià, Girona, Spain (42°10′29.4″N, 3°02′41.0″E). M. Riutort, coll., 2010. F6108: Cártama, Málaga, Spain (36°42′46.6″N, 4°37′53.0″W). E. Mateos, leg., 10/mar/2014. A portion 10 mm long including pharynx, sagittal sections on 32 slides; a portion 10 mm long including copulatory apparatus, sagittal sections on 33 slides. Harvard IZ 59101 (field number F6099): Torroella de Fluvià, Girona, Spain (42°10′29.4″N, 3°02′41.0″E). M. Riutort and F. Carbayo, coll., 19/Mar/2014. Specimen killed with boiling water and fixed in 80% ethanol. Anterior end 5 mm long kept frozen in absolute ethanol, rest of the body in clove oil for 15 days for visualizing distribution of eyes, and preserved in 80% ethanol. NHMW 2743, Series 2: no available locality. Three slides, serial sagittal sections of copulatory apparatus.
Tests on food preferences were conducted with specimens F6101, F6105, F6106, F6100 and F6108.
DESCRIPTION
Extended living worms were 5–7 cm in length, about 5 mm in width and 1–2 mm in height ( Figs 11–14 View Figure 11 View Figure 12 View Figure 13 View Figure 14 ; http:// www.youtube.com/watch?v=sZU_ncoawfY). Contract- ed, they are about 3 cm long and 1.1 cm wide. The anterior end is rounded, narrow, about 1 mm wide and widens to the maximum body width about 1.5 cm from the anterior tip. Thereafter the sides of the body are roughly parallel until about 1 cm from the posterior when it tapers to the pointed tip. On initial viewing of live specimens, the dorsal colour is ‘beige brown’, although some specimens were very dark, almost black. The surface is mucus-covered. On closer examination the dorsal ground colour is ‘golden yellow’ to ‘honey yellow’ with numerous black or ‘terra brown’ spots aligned or aggregated into short longitudinal streaks.
However, there is variation. Some specimens have a narrow pale midline of the ground colour devoid of dark pigment spots. This midline may widen slightly over the regions of the pharynx and copulatory apparatus and may or may not be bordered by a darker region caused by fine patterning of very small dark pigment spots. The very dark, almost black, specimens have a more dense distribution of dark pigment spots almost obliterating the ground colour. The ventral surface is uniformly ‘cream’ or ‘grey white’ and the creeping sole occupies the full ventral width.
Eyes are present in a uniserial row round the anterior tip but polyserial further posteriorly where they spread onto the dorsal surface to the extent of about one-third of the width on either side, and thus about 0.67 of the width ( Figs 15 View Figure 15 , 16 View Figure 16 ).
In preserved specimens, the position of the mouth, on the ventral surface behind the anterior end, varies from 42 to 81% of total body length, median 60.3% ( Table 3 View Table 3 ). The gonopore varies from 59 to 87% of total body length, median 77.1% ( Table 3 View Table 3 ). The sensory pits are simple invaginations 30 μm deep, ventromarginally distributed in a single row externally to the glandular margin, along the anterior third of the body. The epidermis is ciliated over the creeping sole only. Abundant glands with erythrophilic gross granules (1– 2 μm) and a second type containing xanthophilic granules variously sized (1–5 μm) cross the ventromarginal epithelium constituting a glandular margin 100 μm wide.
The three typical geoplaninid cutaneous muscle layers extend throughout the body: one circular followed by a diagonal with deccusate fibres, and then a longitudinal layer with fibres arranged into bundles with 8–25 fibres each, dorsally (20 μm) and ventrally (25 μm) as well. The cutaneous musculature thickness relative to body height at the pre-pharyngeal region (CMI) is 4.6– 6.7%.
The dorso-ventral parenchymal muscle fibres are apparent between the intestinal branches. Three parenchymal muscle layers are present at the pre-pharyngeal region: a dorsal layer of decussate diagonal fibres, a supraintestinal layer of transverse fibres and a subintestinal layer, above the ventral nerve plate, with loose transverse fibres.
The nervous system consists of a broad ventral plate, 70–80 μm in dorso-ventral thickness, extending almost the full width of the animal. There is a slight enlargement to about 100 μm on each side in the vicinity of the ovovitelline ducts.
The mouth is behind the middle of the pharyngeal pouch ( Fig. 17 View Figure 17 ). There is an oesophagus (but it also may be absent as in specimen MZUSP PL 1597, seemingly due to contraction at the time of fixation), 0.6 mm in length, leading to the digestive diverticula. The pharynx is cylindrical with the dorsal insertion slightly posterior to the ventral insertion. It is 2.0–3.4 mm in length, occupying 61–90% of the pharyngeal pouch. The outer epithelium of the pharynx is flat, ciliated, underlain by a layer (10–15 μm) of longitudinal muscle fibres with circular fibres interspersed; the inner pharyngeal epithelium and that of the oesophagous is ciliated; it is underlain by a layer (80–150 μm thick) of circular fibres with longitudinal fibres interspersed.
Glands of three types, producing erythrophilic,
xanthophilic or scarce cyanophilic granules, pierce the
distal portion of the pharynx.
Testes are dorsal to the digestive diverticula but below the supra-intestinal muscle layer, and commence anterior to the ovaries. The most anterior testes are about 1/6th of the body length from the anterior extremity, and extend back to almost the level of the root of the pharynx. Up to six testes may be visible in a transverse section on each side of the body. The testes are spherical or ovate, about 200 μm in dorso-ventral height, 240 μm antero-posteriorly. The paired sperm ducts run posteriorly roughly above the oviducts. About 1 mm from the copulatory apparatus the sperm ducts each broaden and in most specimens they contain densely packed spermatozoa (which are cyanophilic). They turn dorsally towards the sagittal body plane and open to the paired branches of the prostatic vesicle. These branches join and penetrate the antero-dorsal aspect of the penis bulb and continue into the ejaculatory duct, which runs through the centre of the penis to open at its tip ( Figs 18–23 View Figure 18 – 23 ; Figs S18–S36).
The epithelium of the prostatic vesicle is columnar in the proximal portion, 40 μm high, or cuboidal, 15 μm high, in the distal portion. It is pierced by glands containing erythrophilic granules. The lining of the ejaculatory duct is cuboidal and ciliated.
The penis, 1–1.7 mm long, is ovoid-blunt or irregular, and projects posteriorly and to the left from the right dorsal part of the male atrium (but in paratype NHMUK 2013.9.5.1 it is nearly horizontal), not from any atrial fold. Its dorsal insertion is placed nearly in the distal part of the male atrium. The male atrium is an ample cavity mostly occupied by the penis, and provided with a large sheath-like fold on the right side. In most cases this fold arises from the ventral anterior side of the atrium to nearly the dorsal level to the gonopore, thus separating the male from the female atrium. The fold may also be reduced, or emerge from the distal portion of the male atrium, as in specimen NHMUK 2013.9.1.
The epithelium of the penis is 8–20 μm in height and non-ciliated. The stroma of the penis contains two types of glands, one containing erythrophilic (reddish) granules and one containing a second type of erythrophilic granules that stain pinkish (most specimens) or bluish (specimens MZUSP PL 1596 and MZUSP PL 1597, fixed with ethanol) ( Figs 24–26 View Figure 24 View Figure 25 View Figure 26 ; Figs S18–S36). The cell bodies of the two types lie outside the penis bulb. In the proximal region of the penis, the necks of the pinkish glands run in discrete bundles, crossing the central space between the ejaculatory duct and the lining epithelium of the penis. These bundles join in larger, conspicuous bundles (> 60 μm in diameter), even at low magnification, as they approach the subapical height of the penis to discharge their content in distinct patches ( Figs 24–27 View Figure 24 View Figure 25 View Figure 26 View Figure 27 , Figs S25, S28, S35). The glands producing reddish granules run singly or in bundles, the latter running near the ejaculatory duct. They open across the whole epithelium of the penis. A 10–15-μm-thick muscle layer of circular fibres with some longitudinal fibres interspersed underlies the epithelium of the penis, the stroma of which is crossed by numerous muscle fibres orientated in various directions, most of them arranged in bundles that run approximately radially in the vicinity of the ejaculatory duct ( Fig. 26 View Figure 26 ).
The lining epithelium of the male atrium is not ciliated, is cuboidal-to-columnar, apically xanthophilic, but flat in the ventral region. The surface of the large fold facing the penis is crossed by very many glands containing very fine granules. It is difficult to determine their affinity for stain. They seem to be erythrophilic granules immersed in slightly cyanophilic cytoplasm ( Figs 24 View Figure 24 , 26 View Figure 26 , 28 View Figure 28 ); this type of gland also pierces part of the left side of the male atrium, giving rise to an annular secretory surface. The cell bodies of these glands are outside the common muscle coat. Characteristically, and differently from the remaining muscles of the male atrium, the musculature underlying the epithelium of the annular band is constituted by circular muscle fibres arranged in loose bundles, the fibres of which lie one upon the other, giving rise to a kind of parallel fence ( Fig. 26 View Figure 26 ); longitudinal muscle fibres are interspersed with circular ones. Otherwise, the glands and muscle fibres underlying the male atrium are as follows: under the epithelium there is a thin layer of circular muscles, followed by a second one of longitudinal muscles. Glands with erythrophilic granules pierce the epithelium. The epithelium up to 400 μm from the gonopore is ciliated.
The pair of ovaries are about one-fifth of the body length behind the anterior end and lie ventrally immediately above the nerve plate. Each ovary is about 500 μm long and about 300 μm wide. The ovovitelline ducts emerge from the latero-dorsal aspect of the ovaries about half way along its length. In specimen NHMUK 2008.4.1.1 each oviduct has a dense clump of cyanophilic material near the junction with its ovary, which appears to be spermatozoa. Behind the gonopore, the ovovitelline ducts turn dorsally and join in the midline, dorsal to the posterior portion of the female atrium ( Figs 17– 22 View Figure 17 View Figure 18 – 23 , 28–29 View Figure 28 View Figure 29 ). The distal ascending region of the oviducts and the common glandular ovovitelline duct receive the products of erythrophilic glands, presumed to secrete the cocoon casing ( Figs 17–22 View Figure 17 View Figure 18 – 23 , 28– 29 View Figure 28 View Figure 29 ). The common ovovitelline duct runs initially posteriorly then ventrally and anteriorly to communicate with the female atrium. The female atrium is long, somewhat funnel-shaped, with its lumen laterally narrowed by 1–2 longitudinal folds. In specimen MZUSP PL 1597 the atrium is abnormally divided into two chambers. Its lining epithelium, apically erythrophylic, is 25 μm in height in the anterior region and 50 μm in the posterior. The epithelium is ciliated at a distance up to 400 μm behind the gonopore. Except for the epithelium around the genital canal and the ventral one close to the gonopore canal, conspicuous gland cells producing erythrophilic (purple) granules pierce the epithelium. The muscularis of the female atrium consists of a layer (10 μm thick) of decussate fibres. The female atrium is about half the length of the male atrium.
The copulatory apparatus has an anterior–posterior extent of 3.26 ± 0.75 mm ( Table 3 View Table 3 ) from the junction of the sperm ducts to the junction of the oviducts.
Biological note
In Guernsey, specimens were found in several garden centres and plant nurseries beneath flower pots and other objects lying on the soil, where they could be abundant. In Torroella de Fluvià (Girona, Spain) adult specimens were found resting together with hatchlings and empty cocoons in a plastic box sunk into the ground. In lab conditions, a specimen fed on an earthworm when they were placed together in a 50 mL plastic vial. The earthworm had been collected from the same locality. One to three species of snail sampled in green areas in the campus of the University of Barcelona (Av. Diagonal, 643) were individually offered to a total of seven specimens of O. nungara . Specimen F6100 (which died days thereafter in the lab) ate a juvenile snail Cochlicella acuta ( O. F. Müller, 1774) and rejected another one previously offered. Specimen F6101 rejected a juvenile C. acuta and a very juvenile Helicidae . Specimen F6101 also rejected an earthworm sampled in the same area. Specimen F6105 rejected a snail. Specimen F6106 rode on a snail and tried to capture it, but it escaped. An unlabelled specimen ate a frozen piece of a cockroach (L. Leria, pers. comm.). Specimen F6108 was collected from an earthworm farm in Málaga (southern Spain) and sent to Barcelona. In Brazil we found the specimens on a grass field among fishing ponds and at the border of a dirt road in a region covered by Atlantic forest. Obama nungara has been record- ed from 45 departements in France, and is referred to as ‘espece marron plate’ (Justine, weblog and website: https://sites.google.com/site/jljjustine/plathelmintheterrestre-invasif; http://bit.ly/Plathelminthe).
The single specimen found in Italy was collected on 7 November 2012 in Martorano, near Cesena (Forlì- Cesena), Emilia Romagna (44°10′34″N, 12°14′35″E) in ornamental plant pots. It was found by Davide Dradi, passed to Danio Miserocchi who contacted H.D.J. with photographs. It has not been further examined. No further specimens have been found (D. Miserocchi email, 25 July 2014).
A single specimen was found in Pembury, Kent, UK (51°08′46″N, 0°19′39″E), coll. M. Jenner, 23.9.2014. Confirmed by H.D.J. by photos and short video.
DISCUSSION






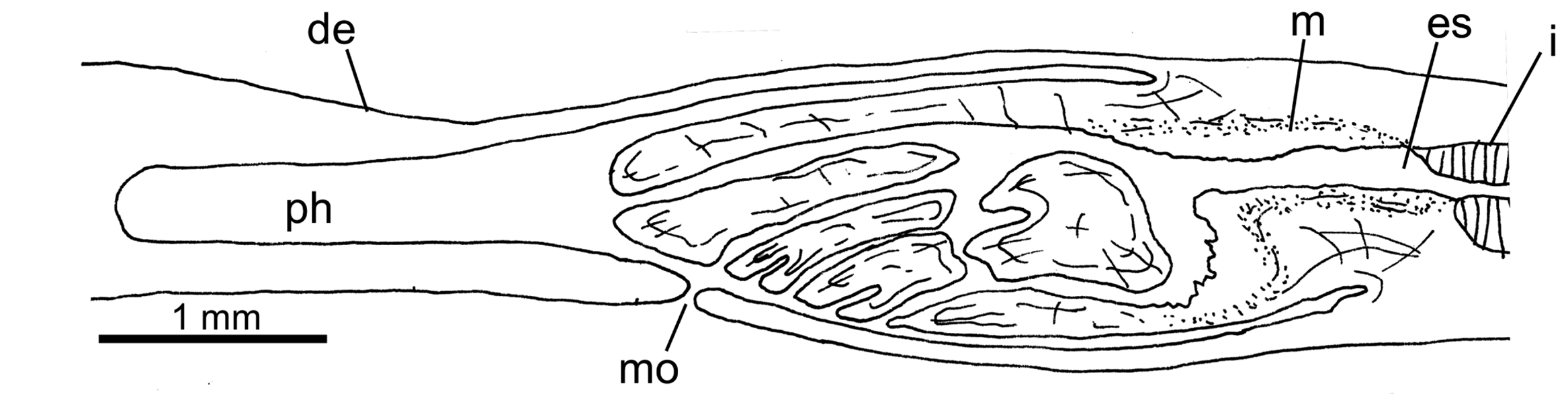
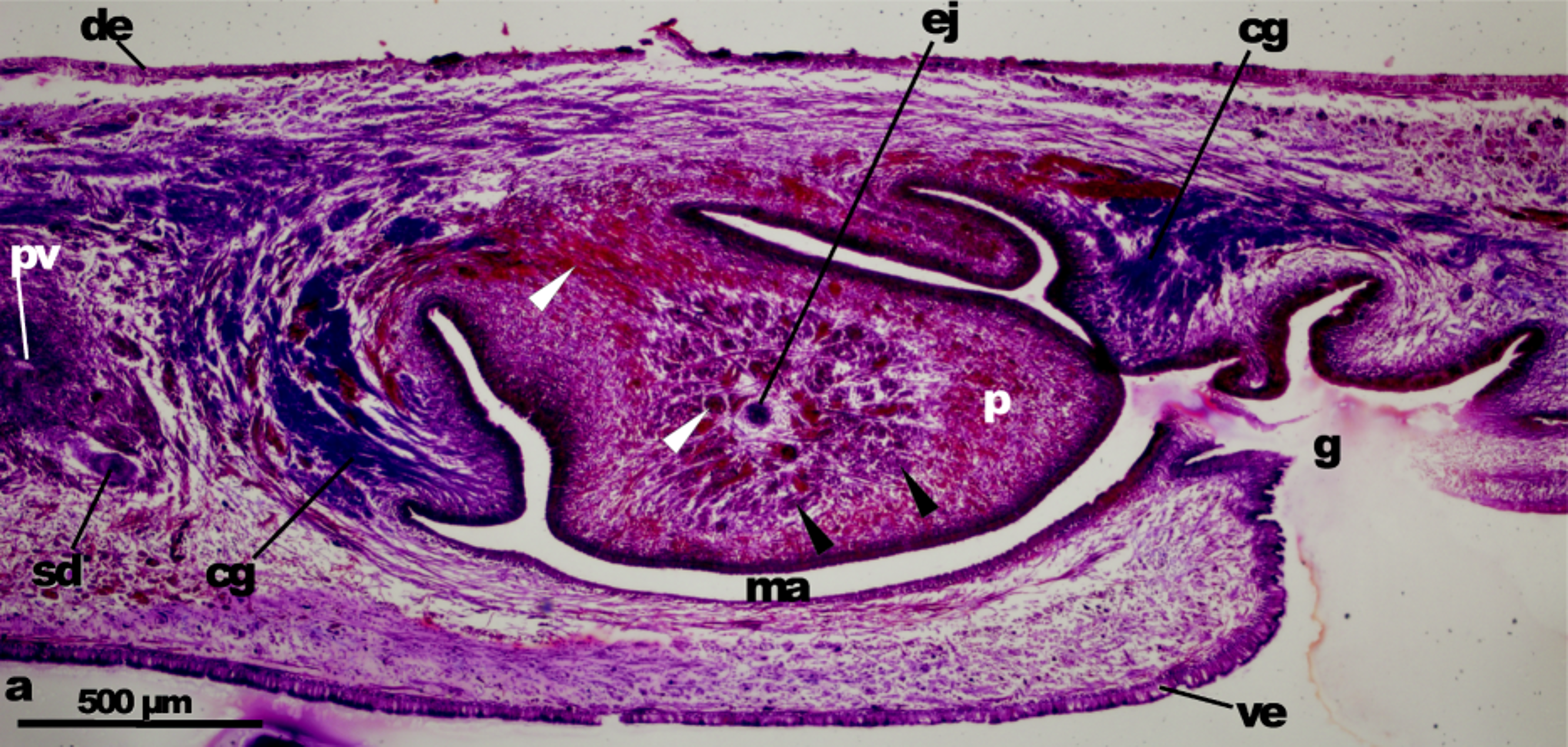





Table 3. Obama nungara sp. nov.: dimensions (mm) of preserved specimens from the UK, Spain and Brazil
| Location | L | W | A-m | A-m% | A-gp | A-gp% | Lcop | |
|---|---|---|---|---|---|---|---|---|
| Guernsey | NHMUK 2008.4.1.1* | 36 | 8 | 19 | 53 | 25 | 69 | 2.3 |
| NHMUK 2008.4.1.2 | 57 | 8.5 | 34 | 60 | 41 | 72 | ||
| NHMUK 2008.4.1.3 | 34 | 6 | 21 | 62 | 26 | 76 | ||
| NHMUK 2008.4.1.4 | 31 | 6 | 19 | 61 | – | – | ||
| NHMUK 2010.10.29.2 | 21 | 4 | 13 | 62 | – | – | ||
| NHMUK 2010.10.29.3 | 39 | 7 | 23 | 59 | 30 | 77 | ||
| NHMUK 2010.10.29.4 | 32 | 7 | 26 | 81 | 28 | 87 | ||
| NHMUK 2010.10.29.5 | 45 | 7 | 27 | 60 | 33 | 73 | ||
| NHMUK 2010.10.29.12 | 33 | 8 | 20 | 61 | 28 | 87 | ||
| NHMUK 2010.10.29.13 | 45 | 8 | 30 | 67 | 38 | 84 | ||
| NHMUK 2010.10.29.14 | 36 | 7 | 25 | 69 | 30 | 83 | ||
| NHMUK 2010.10.29.15 | 40 | 7 | 23 | 57 | 31 | 77 | ||
| NHMUK 2010.10.29.16 | 29 | 5 | 18 | 62 | 23 | 79 | ||
| Fordham | NHMUK 2010.10.29.1 | 62 | 8 | 38 | 61 | 48 | 77 | |
| Spain | NHMUK 2013.9.5.1* | 41 | 7 | 23 | 56 | 32 | 78 | 2.5 |
| NHMUK 2013.9.5.2 | 42 | 6 | 27 | 64 | 33 | 79 | ||
| F6098* | 38 | 6 | 22 | 58 | 28 | 74 | 3 | |
| F6099 | 30 | 7 | 17 | 57 | 23 | 77 | ||
| F6102* | 47 | 6 | 27 | 57 | 37 | 79 | 3.9 | |
| F6108* | 73 | – | 31 | 42.5 | 43 | 59 | 1.9 | |
| F4649* | 38 | 7 | 21 | 55 | 28 | 74 | 3.8 | |
| F4650* | 38 | 7 | 22 | 58 | 29 | 76 | 3.2 | |
| Brazil | F3191* | 48 | 7 | 28 | 58 | 38 | 79 | 3.9 |
| F3247* | 49 | 6 | 29 | 59 | 37 | 76 | 4.1 | |
| F3178 | 44 | 5.5 | 28 | 64 | 34 | 77 | ||
| F3180 | 25 | 4 | 17 | 68 | 20 | 80 | ||
| F3189 | 44 | 4.5 | 25 | 57 | 38 | 86 | ||
| F3190* | 44 | 5 | 27 | 61 | 35 | 80 | 3.8 | |
| F3193 | 31 | 4.5 | 17.5 | 56.5 | 23 | 74 | ||
| F3194 | 49 | 7 | 33 | 67 | 35 | 71 | ||
| F3195 | 38 | 5 | 25 | 66 | 30 | 79 | ||
| F3197 | 29 | 4 | 17 | 59 | 22 | 76 | ||
| F3198 | 34 | 4 | 21 | 62 | 27 | 79 | ||
| F3200 | 25 | 6.5 | 16 | 64 | 19 | 76 | ||
| F3205 | 45 | 6 | 25 | 56 | 33 | 73 | ||
| F3211 | 31 | 4.5 | 19 | 61 | 25 | 81 | ||
| F3212 | 32 | 6 | 19 | 59 | 24 | 75 | ||
| F3227* | 37 | 5 | 215 | 58 | 28 | 76 | 3.5 | |
| F3246 | 25 | 5 | 17 | 68 | 20.5 | 82 | ||
| F3248 | 37 | 4 | 23 | 62 | 29 | 78 | ||
| Median | 60 | 77 | ||||||
| Mean ± SD | 39 ± 10 | 6 ± 1.3 | 23 ± 5.4 | 30 ± 6.7 | 3.3 ± 0.75 |
L, length; W, width; A-m and A-gp are distances from the anterior end to the mouth (pharyngeal aperture) and to the gonopore, respectively. % values are for these as a percentage of total body length. *Sectioned specimens. Lcop, length of the copulatory apparatus from the junction of the sperm ducts to the oviduct-female genital canal.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Obama nungara
| Fernando Carbayo, Marta Álvarez-Presas, Hugh D. Jones & Marta Riutort 2016 |
Obama marmorata
| Lago-Barcia et al. 2015: 17 |