
Parapronoe Claus, 1879
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4192.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:B3AE1A8B-EE40-4ACF-879B-33B55FBD1FB8 |
|
DOI |
https://doi.org/10.5281/zenodo.6069281 |
|
persistent identifier |
https://treatment.plazi.org/id/4A641514-1850-FF9D-FF5E-FB82FCFBFEB2 |
|
treatment provided by |
Plazi |
|
scientific name |
Parapronoe Claus, 1879 |
| status |
|
Genus Parapronoe Claus, 1879
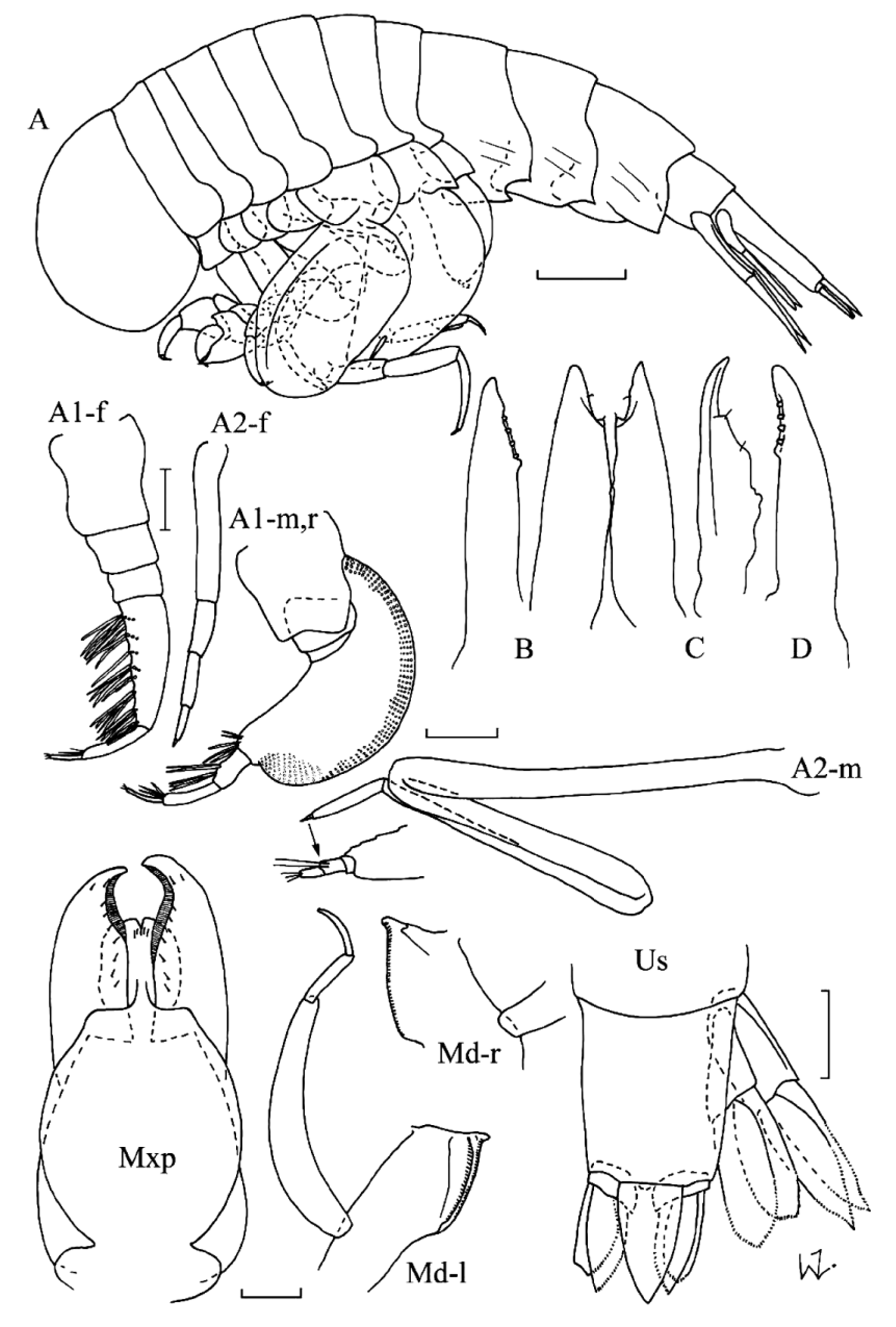
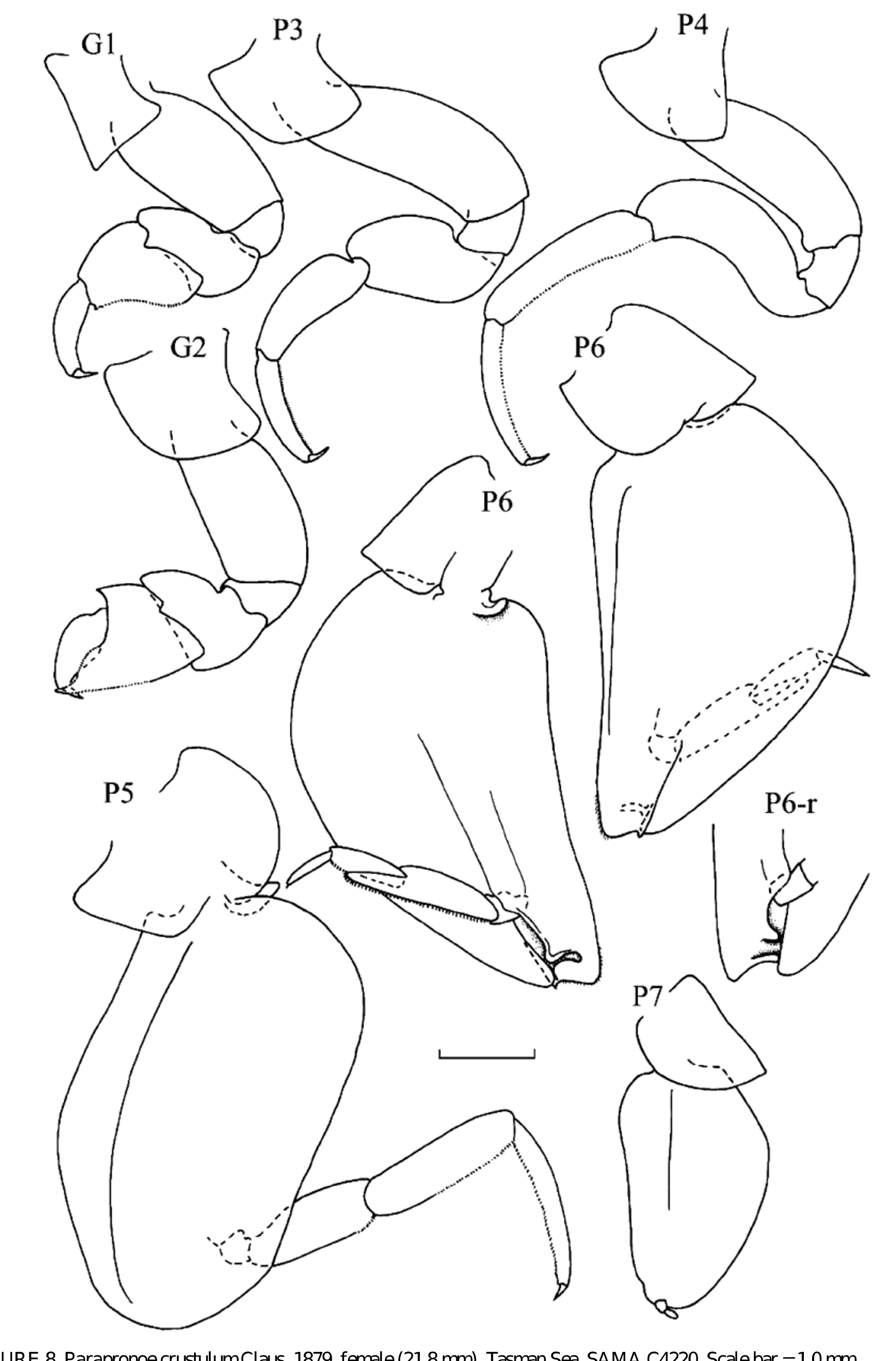
( Figs 7–8 View FIGURE 7 View FIGURE 8 )
Amphipronoe Bate, 1862: 335–336.
Parapronoe Claus, 1879: 23 (key), 29–31.— Gerstaecker 1886: 486.— Claus 1887: 48 (key), 63–64.— Stebbing 1888: 1521 – 1522.— Schellenberg 1927: 651 –652.— Spandl 1927: 217 (key).— Pirlot 1929: 135.— Hurley 1955: 175 (incl. key).— Yoo 1971: 61 (key).— Bowman & Gruner 1973: 42 (key), 43–44.— Zeidler 1978: 25.— Vinogradov et al. 1982: 361 (key), 377– 378.— Shih & Chen 1995: 146 (key), 158.— Vinogradov 1999: 1202 (key), 1203.
Sympronoe Stebbing, 1888: 1533 .— Spandl 1927: 217 (key).— Pirlot 1929: 151.— Bowman & Gruner 1973: 42 (key), 44–45.— Zeidler 1978: 21 (key), 24.
Type species. Parapronoe crustulum Claus, 1979 by page priority. Type material could not be found at the MFN or ZMH and is considered lost. However , the description and figures provided by Claus (1879, 1887) readily characterise this genus. No precise type locality is given, but specimens were collected from the tropical north-east Atlantic , off Lagos and the tropical south-west Indian Ocean, off Zanzibar .
Type species of synonyms. The type species of Amphipronoe is A. cuspidata Bate, 1862 by monotypy . Type material could not be found at the NHM or the MNHN and is considered lost. However, it appears to be a species of Parapronoe , judging by the description and figures provided by Bate, but Amphipronoe has not been used in the scientific literature since Streets (1877) and Giles (1888) (for A. longicornuta a synonym of Lycaea vincentii ), and is rejected as a senior synonym of Parapronoe in order to preserve nomenclatural stability ( ICZN, Article 23.9). The type locality is the South Atlantic, near the Cape of Good Hope, Captain Raynaud , February 1829 .
The type species of Sympronoe is Parapronoe parva Claus, 1879 ; designated by Stebbing (1888). Type material could not be found at the MFN but the ZMH has seven males and one specimen with the head missing ( K22286 View Materials ), which probably represents type material. This material seems to be from the time of Claus and has the locality as “ Sansibar ” ( Zanzibar), which is the type locality provided by Claus. Stebbing (1888) erected this genus for P. parva and a new species, S. propinqua (regarded a synonym of P. parva ), because he believed that they possed characters that distinguished them from Parapronoe . However, Semenova (1981) described a new species of Parapronoe ( P. elongata ), and found that it had features in common with Sympronoe , and concluded that the generic status could not be maintained.
Diagnosis. Body shape robust or globular. Head with short, rounded rostrum in both sexes. Eyes occupying most of head surface; grouped in one field on each side of head. Antennae 1 of males with 3-articulate peduncle; with enlarged, crescent-shaped callynophore, with aesthetascs arranged in one-field brush medially; with three smaller articles inserted on antero-dorsal corner. Antennae 1 of females with 3-articulate peduncle; callynophore narrowly rectangular; with two smaller articles inserted terminally. Antennae 2 of males 6(7)-articulate; strongly zig-zagged, with most articles folded back on each other; only extending anteriorly under head, or also posteriorly between the gnathopoda and pereopoda to pereonite 2; basal article elongate, longer than following article, or about half-length of following article; with small terminal article, not folded, pointing posteriorly, or medially. Antennae 2 of females 5-articulate. Mandibular incisor relatively broad, with several teeth, with small distal lobe medially; in male orientated at right angles to palp. Maxillae 1 consisting of elongate lobes with four rounded teeth distally on medial margin. Maxillae 2 consisting of elongate lobes, curved and pointed distally, with slight medial bulge. Maxilliped with inner lobes incompletely fused, slightly separate terminally; medial margin of outer lobes with fringe of closely packed slender setae. Gnathopod 1 simple. Gnathopod 2 chelate; carpal process knife-shaped, armed with microscopic teeth or setae. Pereopods 3 & 4 distinctly shorter than pereopods 5 & 6. Pereopod 5; basis very broad, more than 5 x as wide as merus, but not operculate, may overlap with P6 and lock proximally; articles 3–7 inserted sub-terminally to basis. Pereopod 6; basis very broad, more than 5 x as wide as merus, but not operculate, may overlap, or lock, with opposing pereopod; articles 3–7 inserted sub-terminally on basis; merus with antero-distal corner extended, distinctly overlapping carpus medially. Pereopod 7 reduced in size with large basis; with only 1–3 terminal articles. Uropoda all with articulated exopoda and endopoda, those of U1 & 2 lanceolate, usually with serrated margins. Uropod 3 with endopod, and sometimes also exopod, leaf-like, foliaceous, with smooth margins.
Species. Parapronoe crustulum Claus, 1879 ; P. parva Claus, 1879 ; P. campbelli Stebbing, 1888 and P. elongata Semenova, 1981 .
Sexual dimorphism. The sexes are very similar in gross morphology, differing mainly in the morphology of the mandibles and the antennae. Females also tend to have a more rounded head than males, and reach a slightly larger size.
Remarks. Parapronoe resembles genera of the families Parascelidae and Platyscelidae in their ability to curl up into a ball. In all species, the coxa of pereopod 5 articulates proximally with the coxa/basis of pereopod 6, and, except for P. campbelli , the basis of pereopod 6 has a strong ridge/groove structure distally, on the medial surface, which allows the opposing pereopods to lock together. This would allow the animal to enclose the pereopods with the broad basis of pereopods 5 and 6. Parapronoe campbelli lacks a locking mechanism on pereopod 6, but there is a thin ridge along the mid-medial surface of the basis, which may allow the animal to hold the pereopods together. In P. crustulum the first epimeral plate has a postero-distal excavation, which allows the pleon to curl up neatly, and the third epimeral plate has a distinct groove, into which fits the basis of pereopod 6. In P. parva the pleonites have a strong lateral ridge which, when the pleon is curled up, lines up, and fits neatly, with the posterior margin of the basis of pereopod 6. In the few specimens of P. elongata that were examined, there was no evidence of any modification of the pleonites in relation to curling. In P. campbelli the third epimeral plate has a slight ridge, but this does not seem to be related to curling, and it is likely that this species, which also lacks the locking mechanism of pereopod 6, is unable to curl up into a ball.
Parapronoe crustulum differs significantly from all congeners in the morphology of the second antennae of males, in that they do not extend posteriorly underneath the pereon, the basal article is the longest, and the fourth article is relatively short, with 2–3 tiny terminal articles, together orientated medially under the head. In all other species the second and third articles are the longest, and extend posteriorly to about pereonite 2, and the fourth article is about one-third the length of the preceding one, with two tiny terminal articles, which together are orientated parallel to the preceding articles.
As part of this study, specimens from the collections of many museums were examined, including type material of Typhis rapax Milne-Edwards, 1830 , held by the MNHN, which proved to be the same as P. crustulum ( Zeidler 1996b) . As this threatened the stability of some well-established scientific names an application to the ICZN was made to conserve the specific name of Parapronoe crustulum Claus, 1879 ( Zeidler 1996a) , which was subsequently upheld by the ICZN (1998).
Virtually nothing is known about the biology of species. Harbison et al. (1977) found P. parva associated with the siphonophore Rosacea cymbiformis.
Parapronoe appears to be widely distributed in tropical and temperate regions of the world’s oceans. Parapronoe elongata is a relatively rare species, and is only known from the Tasman Sea ( Semenova 1981), the Coral Sea ( Zeidler 1998) and the China Sea ( Shih & Chen 1995).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Parapronoe Claus, 1879
| Zeidler, Wolfgang 2016 |
Sympronoe
| Zeidler 1978: 21 |
| Bowman 1973: 42 |
| Pirlot 1929: 151 |
| Spandl 1927: 217 |
| Stebbing 1888: 1533 |
Parapronoe
| Vinogradov 1999: 1202 |
| Shih 1995: 146 |
| Vinogradov 1982: 361 |
| Zeidler 1978: 25 |
| Bowman 1973: 42 |
| Yoo 1971: 61 |
| Hurley 1955: 175 |
| Pirlot 1929: 135 |
| Schellenberg 1927: 651 |
| Spandl 1927: 217 |
| Stebbing 1888: 1521 |
| Claus 1887: 48 |
| Gerstaecker 1886: 486 |
| Claus 1879: 23 |