
Cypris mareotica, Rasouli, Hamidreza, Scharf, Burkhard, Meisch, Claude & Aygen, Cem, 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4154.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:CCA6924F-47B0-4002-B778-DAA590A0CFDD |
|
DOI |
https://doi.org/10.5281/zenodo.6090838 |
|
persistent identifier |
https://treatment.plazi.org/id/4A0587AC-FFF5-FF9B-3485-FF41FBABF86C |
|
treatment provided by |
Plazi |
|
scientific name |
Cypris mareotica |
| status |
|
Eucypris mareotica ( Fischer, 1855)
1855 Cypris mareotica n. sp. Fischer: 653; pl. 19: 39–41.
1903 Cypris inflata n. sp. Sars: 222; pl. 16: 2a–d.
1909 Cypricercus mongolicus n. sp. Daday: 19; figs 5: a–c; 6: a–g.
1910 Non Cypris inflata n. sp. Daday: 165; pl. 9: 9–13. Synonym: Cypris latissima (G.W. Müller, 1898) . 1925 Eucypris inflata nov. comb.—Bronshtein: 11; pl. 1: 9–16.
1928 Eucypris mongolica ( Daday, 1909) nov. comb.—Bronshtein: 85. 1928 Eucypris mareotica ( Fischer, 1855) nov. comb.—Gauthier: 368.
1947 Eucypris inflata ssp. mongolica ( Daday, 1909) —Bronshtein:124; fig. 51: 1–10. Present study Eucypris mareotica ( Fischer, 1855) var. mongolica nov. comb.
Material examined. 1. Iran, temporary saline road side pool covered with aquatic submerged plants located along the Urmia-Tabriz main road ( 37°43’48” N, 45°14’54” E; May 21, 2011, leg. H.R.): c. 65 males and females (sex ratio slightly biased in favor of females). GoogleMaps
2. Iran, saline lake. Hartmann-collection, Zool. Mus., Univ . Hamburg, Germany, leg. Lindberg, det. Gerd Hartmann: 1 adult, micropalaeontological slide number 27586 ; 1 adult, micropalaeontological slide number 27587. Sex unknown.
3. Germany, Holocene sediments from the former Salziger See ( 51°28’17” N, 11°41’37” E): 94 adults and 91 juveniles (recorded as Eucypris sp. in Wennrich 2006) (material examined by B.S.). GoogleMaps
4. Tunisia, a well at Chenini , Palm Garden ( 33°51’52” N, 10°03’20” E, Nov. 11, 2010, leg. B.S.): 3 female adults and 7 juvenile valves. Material studied by B.S. GoogleMaps
5. Astana City, Kazakhstan. Sars-collection, Zool. Inst. St. Petersburg , Russia: 5 females and 9 juveniles. Type material of Cypris inflata Sars, 1903 . Special Journal Number 6789, Sample No. 10: 5 females and 9 juveniles, labeled Cypris inflata s. nova G. O. Sars. Locality: Saline lake at Akmolinsk (present name: Astana City, Tselinograd District, Kazakhstan). Date: June 30, 1899. Material studied by B.S. , June 6, 1995.
6. Greece, eastern coast of Crete, ( 35°15’28” N, 26°15’54” E, April 6, 2009, leg. R. Fuhrmann), 6 males, 10 females, 1 juvenile. Material studied by B.S. GoogleMaps
7. Crimea , 2 samples: (a) Chersonesskoe Lake ( 44°35’10”N, 33°23’32”E, April 26, 2009, leg. E. Nevrova): 13 males, 59 females GoogleMaps . (b) Lake Adjigol ( 45°06’28” N, 35°27’59” E, May 2, 2009, leg. E. Nevrova): 20 males: 54 females. Material studied by B.S. GoogleMaps
8. Greece, 4 samples: (a) coastal lake near Thessaloniki ( 40°28’58”N, 22°49’39”E, Jan. 11, 2013, leg. B.S.) GoogleMaps : 1 male, 16 females, 280 juv. (b) coastal lake near Thessaloniki ( 40°29’02” N, 22°49’39” E, Jan. 11, 2013, leg. B.S.) GoogleMaps : 3 males, 3 females, 59 juv. (c) coastal lake near Thessaloniki ( 40°26’28” N, 22°51’57” E, Jan. 11, 2013, leg. B.S.) GoogleMaps : 1 male, 3 females, 34 juv. (d) coastal lake, Axios delta ( 40°33’31” N, 22°44’29” E, Jan. 13, 2013, leg. B.S.) GoogleMaps : 2 females.
9. China, subrecent samples from Lake Gahai ( 37°07’43”N, 97°33’00”E, Nov. 11, 2010, leg. S. Mischke): 4 adults and 1 juvenile. Sex unknown. Material studied by B.S. GoogleMaps
Abbreviated diagnosis. An ostracod species with subovate carapace in dorsal view. Carapace roundly triangular in lateral view, with marked compression on both valves in postero-ventral area. Valves bright green. Anterior valve surface rough, but without wart-like elevations ( porenwarzen). Valve margins smooth, without denticles or pustules. Natatory setae of A2 well developed, distinctly exceeding tips of terminal claws. Distal segment of Mx-palp cylindrical; both teeth bristles on third endite markedly serrated. Long d1-seta conspicuously present on basal segment of T2, d2-seta missing. Long terminal claw of UR longer than half L of anterior margin of ramus.
Female. Carapace roundly triangular in lateral view, subovate in dorsal view, with maximum W approximately equal to maximum H or slightly smaller. Both valves laterally distinctly compressed in postero-ventral area ( Fig. 2 View FIGURE 2 A). LV overlaps RV along the entire valve margins ( Figs 2 View FIGURE 2 A, C, D). Fused eye and ovaries clearly visible through carapace.
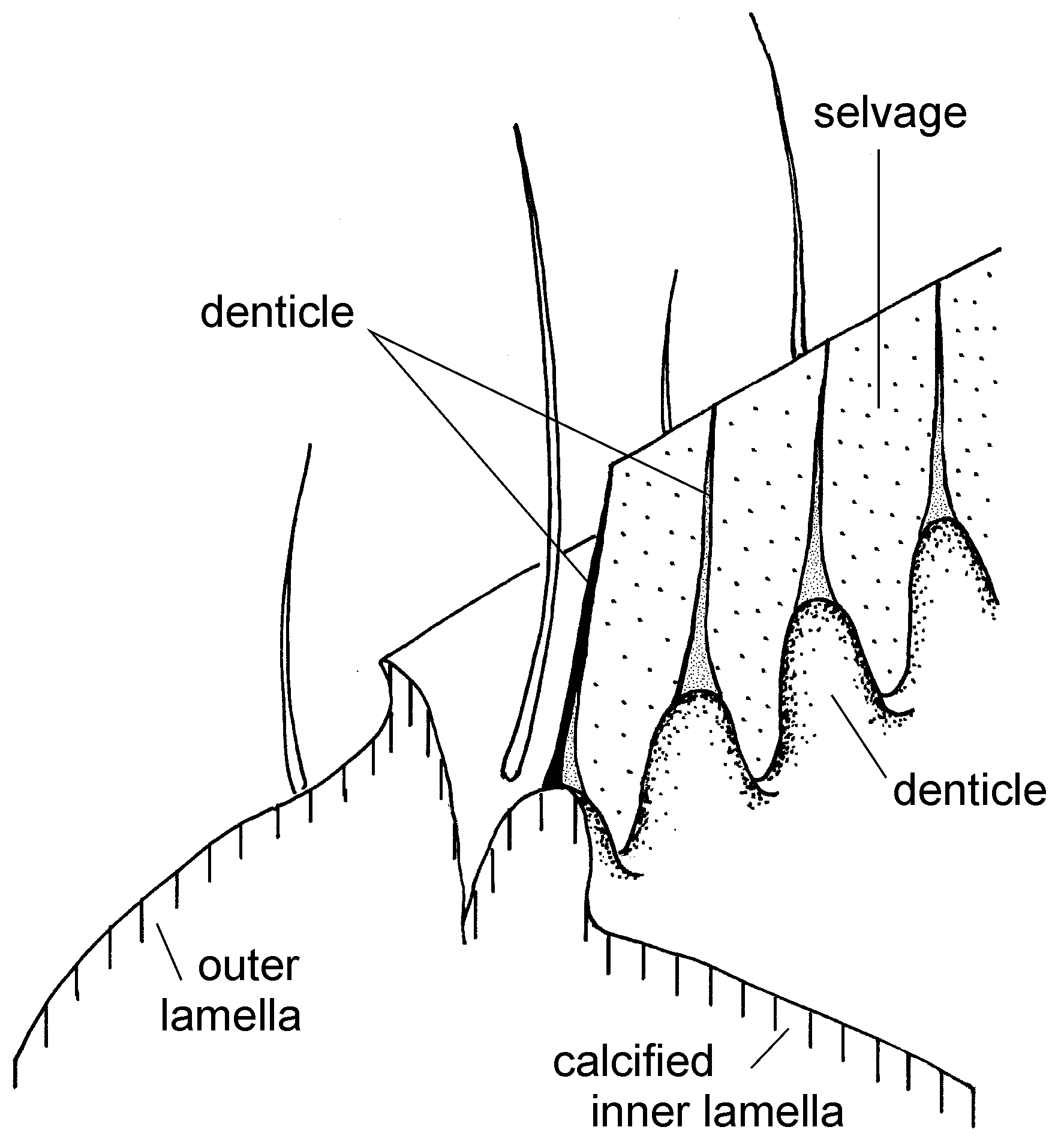
Valve surface with sparse, short and delicate setae, lacking distinct wart-shaped elevations ( porenwarzen) in anterior marginal valve area. Both valves anteriorly and postero-ventrally with inner reinforcements mimicking tiny denticles (only seen in transmitted light at more than 300x magnification) ( Fig. 3 View FIGURE 3 ). L: 1.12–1.17 mm (average 1.15 mm; n=9).
LV ( Fig. 2 View FIGURE 2 E, I, K) roundly triangular in lateral view, maximum H situated in front of mid-length. Dorsal margin evenly joining to anterior end, but with round angle towards posterior end. Anterior margin broader than posterior one. CIL anteriorly broader than posteriorly. Valve margins entirely smooth, without denticles or pustules. H: L ratio c. 0.63.
RV ( Fig. 2 View FIGURE 2 F, J, L) similar to LV, but slightly smaller. Ventral concavity at mid-length more developed than of LV.
A1 (Fig. 4A) 7-segmented, basal segment ventrally with two unequally long setae and one shorter seta dorsally. Second segment with one seta reaching end of next segment; Rome organ present. Wouters’ organ not seen. Third segment with two apical setae, one of which exceeds tip of distal segment and shorter seta reaching to end of next segment. Fourth segment dorsally with two very long setae, this segment additionally with shorter seta reaching utmost to middle of sixth segment and longer seta reaching beyond tip of terminal segment. Fifth segment with two groups of setae: dorsally with two very long and ventrally with two short setae (three times as long as combined sixth and seventh segments). Sixth segment apically carrying four very long setae as well as α-seta about 3.5 times as long as terminal segment. Terminal segment with two very long setae, aesthetasc ya and one thin seta about 2 times as long as ya.
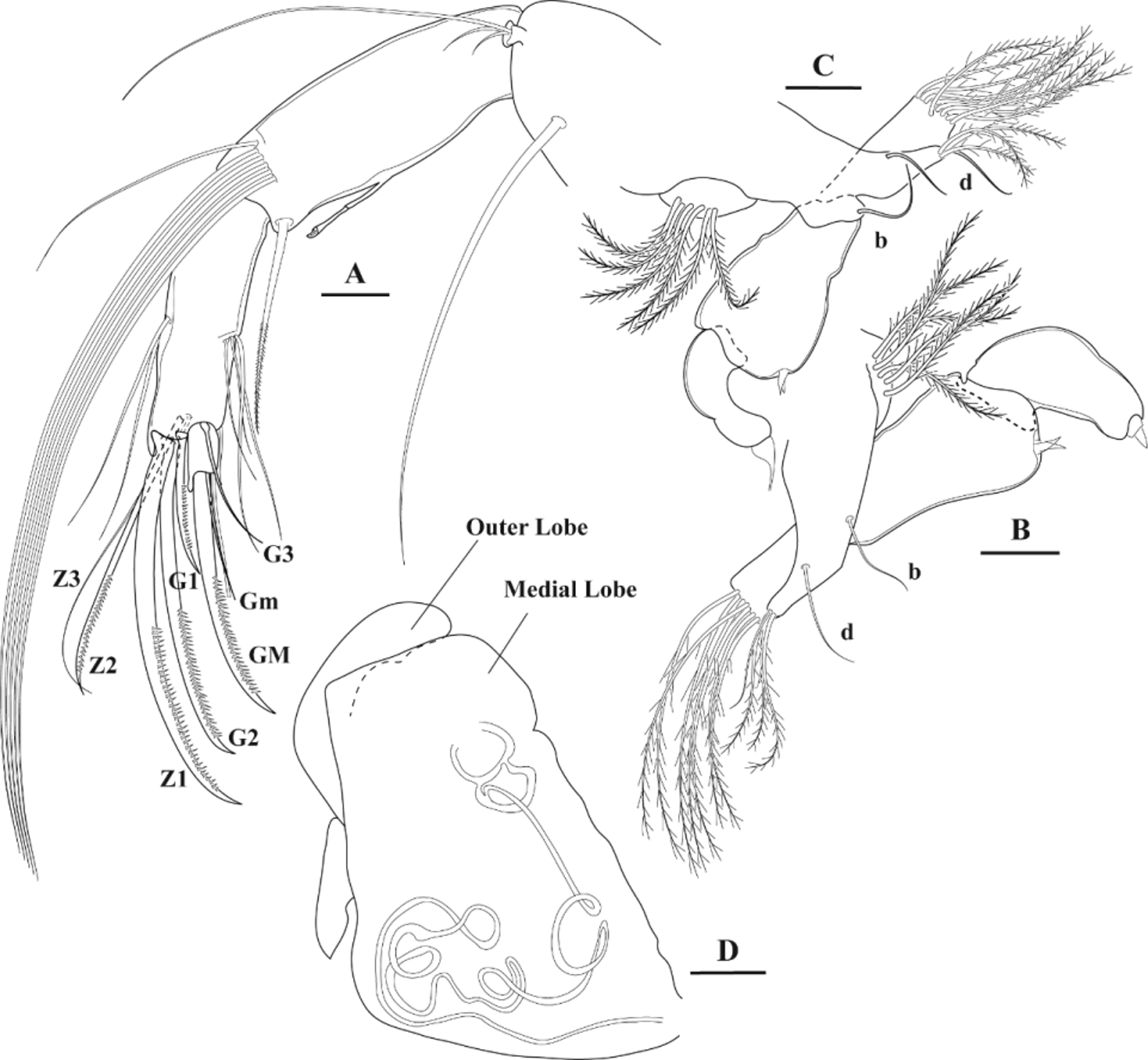
A2 (Fig. 4B). Protopodite: basal segment ventrally with two setae; distal segment with one very long seta and exopodite with three setae (one long and two shorter setae). Second segment of endopodite with 5+1 well developed natatory setae, setae 1–5 distinctly extending beyond tips of terminal claws. Aesthetasc Y well developed. Penultimate segment (third segment) with four unequally developed t-setae and two dorsal setae. Terminal claws G1, G2 and G3 serrated, G1 being longest. Three z-setae present, one of them reinforced, clawlike. Terminal segment with two serrated claws ( GM and Gm).
Md palp (Fig. 4C): Basal segment with two long pappose setae (S1 and S2), one simple seta and one α-seta. Second segment ventrally with group of three equally long setae, shorter plumose seta and stout plumose β-seta; this segment baso-dorsally with three setae. Third segment ventrally with one long and one small seta; dorsal margin basally with four long setae; apically, there are three long setae and one long plumose γ-seta, latter one 1.5 times longer than terminal segment. Terminal segment with three long and two short claws.
Mx (Fig. 4D): Palp 2-segmented; first segment bears six subequal setae (two of them plumose). Terminal segment elongated and cylindrical (about twice as long as broad), with three weakly developed claws and two stout setae. Both teeth bristles on third endite serrated. Vibratory plate with 15 plumose setae.
T1 (Fig. 4E) with protopodite bearing about 12 plumose setae on tip and two long plumose setae (b and d). Two a-setae (both barely visible) and b- and d-setae present, c-seta absent. Palp with three apical setae. Vibratory plate with 5–6 filaments (five filaments in the five individuals from Greece and Crimea, six filaments in one specimen from Greece (collection of B.S.), six filaments in species from Iran (collection of H.R.).
T2 (Figs 4F and 8): Basal segment with very long d1 seta; d2 seta absent. Second and third segments each with one simple seta (setae e and f). Fourth segment with g-seta accompanied with thin additional seta. Fifth segment smallest of all, carrying a strong claw (h2) along with two relatively long setae (h1 and h3).
T3 (Fig. 4G): 4-segmented, distally with pincer organ. Basal segment with d1, d2 and dp setae, dp longest seta of entire limb, longer than L of second segment; plumose e- and f-setae present. Pincer organ with two curved modified setae (h1 and h2) and one long, simple seta (h3).
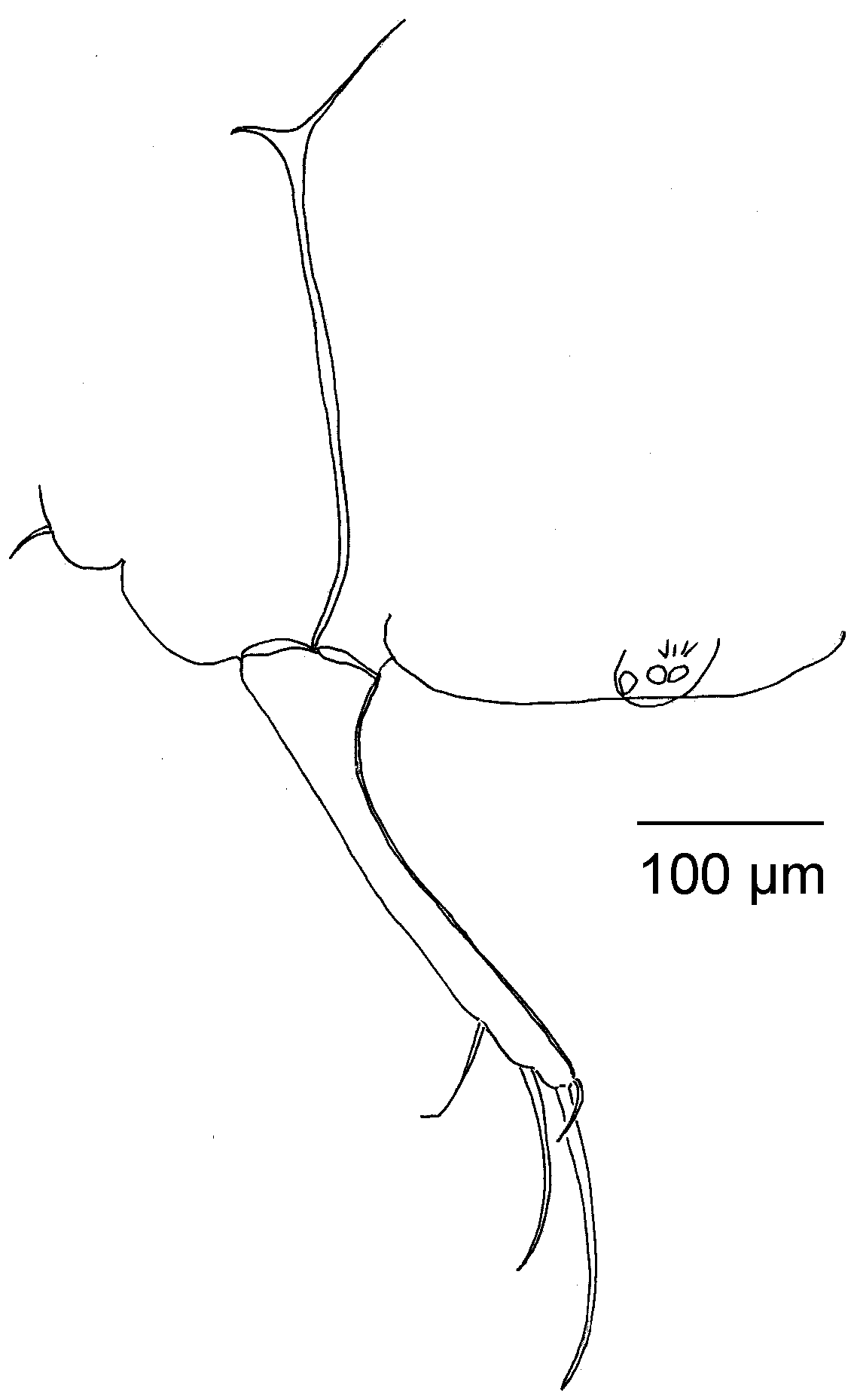
UR (Figs 4H and 5) with cylindrical structure. L ratio of Ga: Gp: Sp: Sa is 1: 0.77: 0.53: 0.21. Attachment of UR proximally simply branched.
Genital lobe ( Fig. 5 View FIGURE 5 ) flat, without any process.
Male. Carapace similar in shape and structure to that of female but shorter ( Fig. 2 View FIGURE 2 B, G, H, M, N). L = 0.98– 1.01 mm (average 1 mm, n = 8). Testes clearly visible through valves. Soft parts similar to those of female except:
A2 ( Fig. 6 View FIGURE 6 A): G3 claw on penultimate segment reduced, seta-like. G1 claw finely serrated, reduced to about 1/ 3 the L of G2 claw. z1 and z2 setae modified into serrated and stout claws, z3 relatively long seta. Gm-claw on apical segment weakly developed in comparison to that of female. In general, A2 of male somewhat longer and more elongate than of female.
T1 ( Fig. 6 View FIGURE 6 B, C): Right and left prehensile palps asymmetrically built, but both endites similar to those of female. Right prehensile palp ( Fig. 6 View FIGURE 6 B) large and stout, left one distally hook-shaped ( Fig. 6 View FIGURE 6 C).
UR: L ratios of Ga: Gp: Sp: Sa = 1: 0.71: 0.56: 0.19.
Copulatory organ ( Figs 6 View FIGURE 6 D and 7): Medial lobe subrectangular; outer distal lobe long and broad, distally recurved, proximally with narrow expansion.
Zenker organ ( Fig. 9 View FIGURE 9 ) with c. 30–32 rings of spines.
Remarks and affinities. Fischer (1855) firstly and very briefly described Eucypris mareotica (as Cypris mareotica ) from Mareotis Lake (today Lake Mariut), a coastal brackish water lake in the vicinity of Alexandria (Egypt). No type material of the species described as new by S. Fischer is expected to exist today ( Damkaer & Matzke-Karasz 2012) and therefore Fischer’s material of E. mareotica has to be considered as lost. The second author (B.S.) travelled to Alexandria to collect ostracods from Lake Mariut with the intention to propose a neotype of the species, based on specimens collected from the type locality. However, Lake Mariut is heavily contaminated by sewage water and has reduced salt content (or has lesser salinity) at present, which probably explains why E. mareotica could not be found at the type locality.
Eucypris inflata ( Sars, 1903) is generally considered a synonym of E. mareotica ( Bronshtein 1947, Martens 1984, Baltanás et al. 1990, Martens & Savatenalinton 2011, Karanovic 2012), and this synonymy is also adopted here. Eucypris inflata was originally described as Cypris inflata by Sars (1903) from the saline lake Djar-Sor in Kazakhstan. The valves of the type specimens are presently decalcified but the shape of the carapace is recognizable. During the study of the type material, the second author has dissected two adult females on two slides for permanent preservation and drawn the appendages (e. g., Fig. 8). Both the carapaces and appendages of the type specimens are clearly congruent with the remaining material studied. Sars (1903), Gauthier (1928: figs 1A–C), Bronshtein (1928, 1947: fig. 50) and Hartmann (1964) mentioned the existence of tiny denticles along the valve margins of Eucypris inflata . We here confirm the existence of these denticles ( Fig. 3 View FIGURE 3 ), which, in fact, are internal reinforcements emerging from the inner calcified lamella to penetrate into the selvage. Due to the internal structures, these denticles are not seen in the SEM images ( Fig. 2 View FIGURE 2 ; see also the SEM images provided by Fuhrmann & Goth 2011 and Fuhrmann 2012). Based on the present knowledge, these structures have so far not been described from any other cypridid ostracod. Fuhrmann & Goth (2011: 136; pl. 9: 10–13) mentioned the presence of very low porenwarzen on both valves, situated close to the anterior margin as well as in the postero-ventral compressed valve area. Although it is rough in those areas, the valve surface of our specimens does not exhibit those structures.
FIGURE 4. Eucypris mareotica ( Fischer, 1855) . Female: A, A1; B, A2; C, Md palp; D, Mx; E, T1; F, T2; G, T3; H, UR. Scales: 43 µm.
Eucypris mareotica is generally assigned to the genus Eucypris View in CoL within the subfamily Eucypridinae Bronshtein, 1947 View in CoL . Bronshtein (1928) noted that the male clasping organs and the uropodal rami of E. mareotica were similar to those of Cyprinotus salinus View in CoL (today Heterocypris salina View in CoL ). Gauthier (1928) wrote that, among other characters, the ornamentation of the selvage, the shape of the male clasping organs and the structure of the hemipenis placed E. mareotica somewhat apart from Eucypris View in CoL s.s.
According to Martens & Savatenalinton (2011) and Petkovski et al. (2016), the subfamily Eucypridinae View in CoL , with the single tribe Eucypridini, comprise eight and ten genera, respectively. The most important differential feature of the members of the Eucypridinae View in CoL is the presence of the so-called c-seta on T1 (maxilliped). According to Meisch (2000) the members of this genus are additionally characterized by smooth (not barbed) teeth bristles on the third endite of the Mx (with the exception of the genera Trajancypris and Arctocypris View in CoL ) and the presence of two setae (d1 and d2) on the basal segment of T2. Eucypris mareotica does not comply with that diagnosis. We do not have seen a c-seta on the T1 of the type material, including the specimens dissected by ourselves (Fig. 4E); both Mx-teeth bristles are distinctly serrated (Fig. 4D) and there is only one d-seta on T2 (d1; Figs 4F and 8). Furthermore, the penis of E. mareotica differs from that of Eucypris virens View in CoL , the type species of the genus, as follows: the outer distal lobe is longer and distally recurved, and additionally, possesses what appears to be a rather conspicuous proximal expansion (outer distal lobe shorter and without a proximal expansion in E. virens View in CoL ). From all this it becomes clear that E. mareotica has to be removed from Eucypris View in CoL sensu stricto. However, as no other genus is available at the present state of knowledge to better accommodate this species, we provisionally prefer to refer to it as Eucypris mareotica . Interestingly, the penis of Eucypris pigra (Fischer, 1851) bears an outer distal lobe-expansion similar to that of E. mareotica . It is well known that E. pigra has probably to be removed from Eucypris ( Meisch 2000: 294) View in CoL . However, as E. pigra differs in other important taxonomic features (see Meisch, op. cit.), no close phylogenetic relationship of E. pigra and E. mareotica is suggested here. A very similar expansion of the outer distal lobe is also seen in the penis of Eucypris persica Klie, 1937 . Klie (1937: 210) expressively draws the attention to the similarity of the penes of E. persica and E. inflata . Therefore, we suggest that E. persica has probably to be removed from the genus Eucypris View in CoL .
Eucypris mongolica ( Daday, 1909) Bronshtein, 1928 , firstly described from lake Issyk Kul in Kyrgyzstan and so far only known from there, differs from E. inflata exclusively in the shape of its carapace, which appears more compressed in dorsal view. Bronshtein (1947) concluded that E. mongolica was conspecific with E. inflata , but kept it as a subspecies of the latter: E. inflata ssp. mongolica . In our opinion, the mongolica -form most probably ranks within the variability range of E. mareotica , based on its wide geographical distribution and also the existence of both bisexual and parthenogenetic populations, which are expected to present some variation in carapace shape. We therefore consider the mongolica- form a simple variety of E. mareotica without any taxonomic status: E. mareotica var. mongolica nov. comb.
Eucypris cf. inflata recorded from Mount Carmel in Israel by Martens et al. (2002) most probably does not belong to E. inflata (here synonymized with E. mareotica ), as shown by the presence of two setae, d1 and d2, on the basal segment of the walking leg ( Fig. 1 View FIGURE 1 H in Martens et al. 2002) and the relative large size (c. 2.3 mm for Eucypris cf. inflata and c. 1.0– 1.5 mm for E. mareotica ). The Mount Carmel specimens probably belong to a species close to or identical with E. virens View in CoL .
Ecology. Eucypris mareotica is considered a polyhalophilic species ( Fuhrmann 2012), commonly found within salinity ranges of 12–80 ppt and up to 110 ppt ( Löffler 1961, Baltanás et al. 1990, Santamaria et al. 1992, Mezquita et al. 2005).
Distribution. Eucypris mareotica has been reported from North Africa ( Algeria, Tunisia, Egypt), Greece, Turkey, Iran, central Asia, and China ( Tibet). In Europe it is so far only known from Spain ( Baltanás et al. 1990, Mezquita et al. 2005) and from the fossil record (Holocene) from Salziger See, Germany ( Wennrich 2006). In Iran, E. mareotica is known from a number of both, freshwater and saline habitats: Northern Iran ( Bronshtein 1947), Hamadan (a pool in a hotel), Babol, Bandar Gaz (a swamp between the town and the Caspian Sea), Ramsar (oxbow lake) and Lahijan ( Hartmann 1964), and our record from the athalassic Lake Urmia.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cypris mareotica
| Rasouli, Hamidreza, Scharf, Burkhard, Meisch, Claude & Aygen, Cem 2016 |
Eucypris (
| Meisch 2000: 294 |
Eucypridinae
| Bronshtein 1947 |
Eucypris persica
| Klie 1937 |
Eucypris mongolica ( Daday, 1909 ) Bronshtein, 1928
| (Daday, 1909) Bronshtein 1928 |
Eucypris mongolica (
| Daday 1909 |
Eucypris inflata ssp. mongolica (
| Daday 1909 |
Eucypris inflata (
| Sars 1903 |
Eucypris mareotica (
| Fischer 1855 |
Eucypris mareotica (
| Fischer 1855 |
Eucypris mareotica
| Fischer 1855 |
Eucypris mareotica
| Fischer 1855 |
Eucypris mareotica
| Fischer 1855 |
Eucypris pigra
| Fischer 1851 |