
Tibiagomphus noval (Rodrigues Capítulo, 1985), Rodrigues Capitulo, 1985
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4078.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:3D0D79C2-2583-47F4-BC01-5CF347614B44 |
|
DOI |
https://doi.org/10.5281/zenodo.6077755 |
|
persistent identifier |
https://treatment.plazi.org/id/495E274C-FFDA-FF91-FF43-FA9CECE2F945 |
|
treatment provided by |
Plazi |
|
scientific name |
Tibiagomphus noval (Rodrigues Capítulo, 1985) |
| status |
|
Tibiagomphus noval (Rodrigues Capítulo, 1985) View in CoL
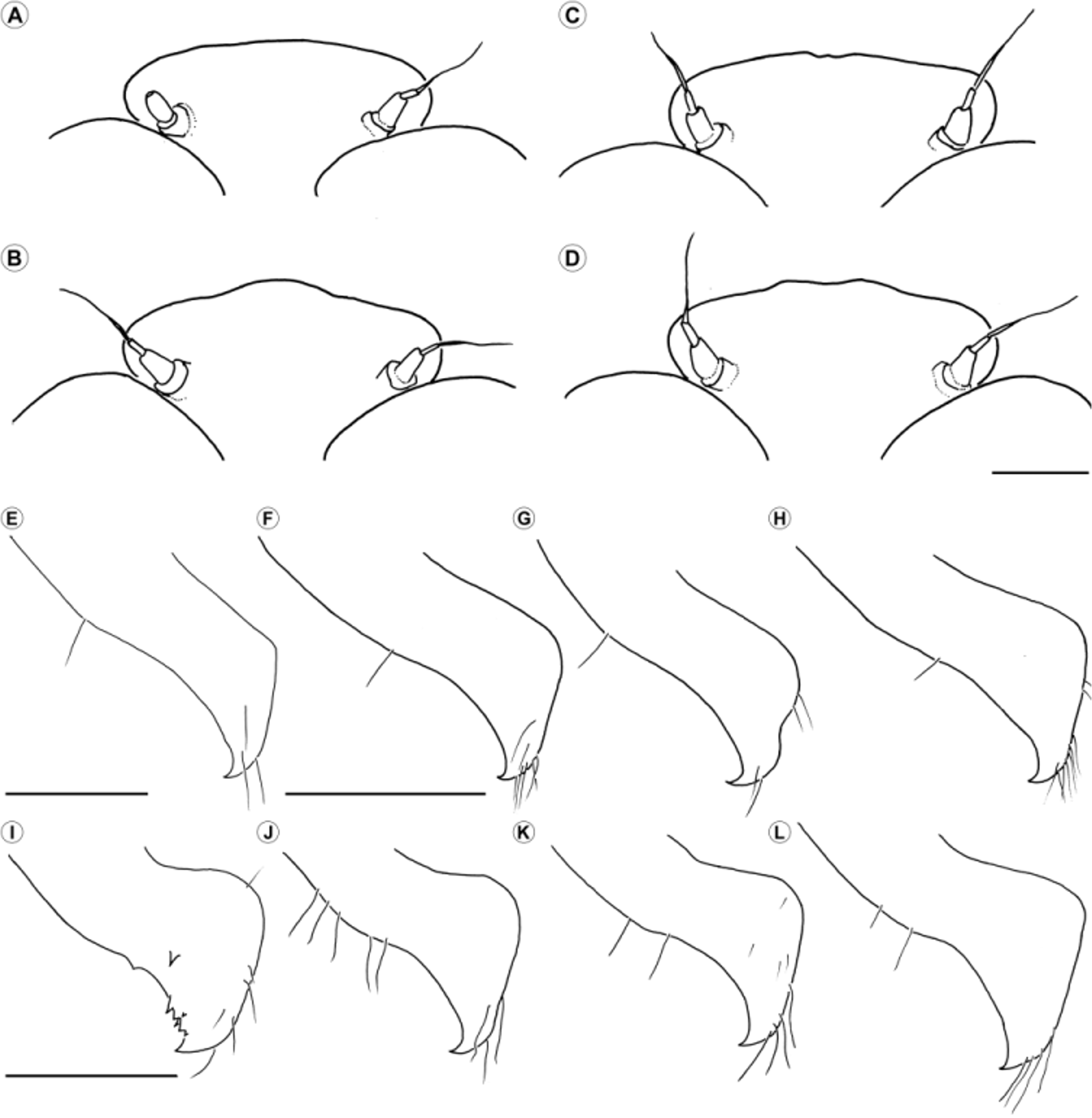
LSID urn:lsid:zoobank.org:act:645A61F2-F59F-42A7-8B73-4270D67477AB ( Figures 2 View FIGURE 2 E, 3C, 4E, 5A–B, F–H, 6F–G, 7J–L, 8G–H, 9E, 10E, 11D, 13)
Cyanogomphus uncatus View in CoL nec Fraser, 1947:—St. Quentin (1973: 358, record from Rio Grande do Sul State, Brazil, likely pertains to this species; comparison with the holotype of T. uncatus View in CoL );— Belle (1980: 153–155, figs 5–13, in part, records from Entre Rios Province, Argentina, and Florida and Durazno Departments, Uruguay, illustrations of metathoracic tibia, tarsi and pretarsal claws in lateral view, V 1–4 in lateral view, V 4 in ventral view, ligula ‘penis guard’ in anteroposterior and lateral views, anterior hamules in ventral view and secondary genitalia in lateral view, comparison with male holotype and female and with C. waltheri View in CoL );— Belle (1981: 261–262, mention);— De Abenante & Philippi (1982: 151, record from Uruguay);— Rodrigues Capítulo (1992: 58, in part, record from Entre Rios Province, Argentina).
Cyanogomphus noval Rodrigues Capítulo, 1985: 329 View in CoL –335, figs 2–14 (description of ♂ holotype, ARGENTINA. Entre Rios Province: Colón Department, Parque Nacional El Palmar, XI.1982, Rodrigues Capítulo leg. in ILPLA [sic MLP], ♀ and F- 0 larva, illustrations of postfrons in dorsal view, secondary genitalia in lateral and ventral views, caudal appendages in dorsal, lateral and ventral views, S 10 in dorsal view of ♂ holotype, subgenital plate and caudal appendages of ♀ in ventral view, habitus of F-0 larvae in dorsal view, head and labium in dorsal view, caudal appendages in dorsolateral view, dorsal margin of abdomen in lateral view comparison with T. uncatus View in CoL and A. ericae View in CoL );— Santos (1988: 267, 286, mention);— Rodrigues Capítulo (1992: 58, 67, 81, 83, 89, figs 1–2, 4–5, 110, 132, 134, 180–181, distribution, reproduction of illustrations from Rodrigues Capítulo 1985);— Garrison (1991: 17, mention).
Tibiagomphus noval (Rodrigues Capítulo, 1985) View in CoL :— Belle (1992a: 1, comb. nov., implicit by the context ICZN 1999, Art. 11.9.3 of the Code);— Belle (1993: 406, mention);— Bridges (1994: VII.168, VIII.70, mention);—Muzón & von Ellenrieder (1999: 125, distributional data);— Garrison et al. (2006: 129–130, comparison, suggest synonym with T. uncatus View in CoL );— Heckman (2006: 525, 534, 649, figs 3.2.589, 3.2.603, key, reproduction of illustrations from Rodrigues Capítulo 1985);— von Ellenrieder & Muzón (2008: 61, record from Entre Rios Province, Argentina);—Muzón et al. (2007: 302, 305, notes on the type material);— Hämäläinen (2015: 114, etymology).
Tibiagomphus uncatus View in CoL nec ( Fraser, 1947):— Belle (1996: 323, fig. 8, in part, pair of wings of ♂ from Uruguay);— Heckman (2006: 649–650, fig. 3.2.778, in part, reproduction of illustrations from Belle 1980 of ♂ from Entre Rios Province, Argentina, and Florida Department, Uruguay);— Garrison et al. (2006: 71, 81–83, 129–130, 329, 331, 337, figs 727, 729a,b, 730, 732, in part, illustrations of mesepisternum in dorsal view and subgenital plate in ventral view of ♀ from Entre Rios Province, Argentina, metathoracic tibia, and secondary genitalia in lateral view from ♂ of Florida Department, Uruguay);—von Ellenrieder & Muzón (2008: 61, in part, record to Entre Rios Province, Argentina);—von Ellenrieder et al. (2009: 229, record from Uruguay);—von Ellenrieder & Garrison (2009: 123, fig. 275, reproduction of illustration of ♀ subgenital plate from Garrison et al. 2006).
Material examined (8♂, 1♀). BRAZIL. Rio Grande do Sul State: 1♂, [Guaíba municipality], Arroio [stream] at BR 116 Km 310 [30°11’45.30”S, 51°23’56.56”W, 20 m a.s.l.], 17.I. [19]79, J.M. Costa leg. ( MNRJ); 1♂, Pelotas municipality [31°46’19.20”S, 52°20’34.08”W, 14 m a.s.l.], 09.III. [19]51, C. Biezanko leg. ( MNRJ 22082); 6♂, Jaguarão municipality [32°33’57.06”S, 53°22’33.60”W, 19 m a.s.l.], Stream at 15 km from highway, 12.XII.1978, J.M. Costa leg. ( MNRJ); 1♀, same data but 12.XIII [sic XII].1978 ( MNRJ).
Type repository. Holotype ♂ by original designation (see remarks) at Museo de La Plata, Universidad Nacional de La Plata, La Plata, Buenos Aires, Argentina ( MLP, examined by photo).
Measurements. Males (n = 8). Total length (including caudal appendages) 45.4–47.0; abdomen length (excluding caudal appendages) 30.8–33.9; head maximum width 5.8–6.3; Fw length 25.8–30.5; Hw length 24.7–26.3; Fw maximum width 4.9–5.5, in Hw 6.1–7.0; pt length 2.6–3.3 in Fw, 3.0– 3.6 in Hw; length of metathoracic femur 7.4–8.2; metathoracic tibia 5.7–6.2; length of S9+ 10 in lateral view 3.2–3.8; total length of cercus in lateral view 1.6–1.9. Ratio cercus length / maximum width 0.30–0.36.
Females (n = 1). Total length (including caudal appendages) 45.2; abdomen length (excluding caudal appendages) 31.8; head maximum width 6.3; Fw length 28.0; Hw length 27.1; Fw maximum width 5.6, in Hw 7.3; pt length 3.1 in Fw, 3.3 in Hw; length of metathoracic femur 8.1; metathoracic tibia 6.2; length of S9+ 10 in lateral view 2.7; total length of cercus in lateral view 1.3.
Larva. Rodrigues Capítulo (1985, p. 332) based on specimens reared in laboratory.
Diagnosis. This pale species, as well as T. uncatus , can be separated from Cyanogomphus spp. except C. comparabilis by the mesepisternum close to the dorsal carina largely pale (dark in C. angelomachadoi sp. nov. and C. waltheri , Figs 1 View FIGURE 1 , 10 View FIGURE 10 A–B, D, 11A, C). The narrow cornet-like distal portion of V 4 in ventral view ( Fig. 6 View FIGURE 6 G, maximum width <0.6 of V3 width vs.> 0.75 in Cyanogomphus , Figs 6 View FIGURE 6 B, E), blade-like cercus distinctly flattened with apex directed posteriorly ( Figs 7 View FIGURE 7 J–L, 8G–H, cercus cylindrical, curved toward to the upturned tapered apex in Cyanogomphus , Figs 7 View FIGURE 7 A–I, 8A–F), the dorsally smooth epiproct, lacking a distal concavity on the epiproct ( Figs 8 View FIGURE 8 H, epiproct with a distal concavity in Cyanogomphus , Figs 8 View FIGURE 8 B, D, F) of males, and subtriangular, long subgenital plate (length> 0.5 of S9) with a narrow and deep mesal incision ( Fig. 9 View FIGURE 9 E, subgenital plate short, length <0.4 of S9, not incised, with a wide V- or U-shaped posteromesal concavity in Cyanogomphus , Figs 9 View FIGURE 9 A–D) of females, separates both species of Tibiagomphus from all Cyanogomphus .
Both sexes of T. noval can be distinguished from it sibling congener T. uncatus by its paler generally yellowish color ( Figs 10 View FIGURE 10 E, 11D, distinctly darker in T. uncatus , Fig. 10 View FIGURE 10 F), and by dorsally pale metathoracic tibia and tarsus ( Fig. 3 View FIGURE 3 C, dark in T. uncatus , Fig. 3 View FIGURE 3 D). The posterior hamule varies greatly, being longer and slender with the “heel” distinctly less pronounced in T. noval ( Figs 4 View FIGURE 4 E, 5F–H, shorter and stouter in T. uncatus with a more pronounced “heel”, Figs 4 View FIGURE 4 F, 5I –L), male cercus robust ( Figs 7 View FIGURE 7 G, 8G–H, ratio cercus length / maximum width 0.30–0.36 vs. more slender, 0.25–0.30 in T. uncatus , Figs 7 View FIGURE 7 M, 8I –J). Further, the smaller size of T. noval (male Hw length 24.7–26.3 vs. 26.8–29.7 of T. uncatus ) and allopatric distribution, with T. noval occurring in biogeographical Pampean province of Chacoan dominion and T. uncatus in Araucaria and Parana Forest provinces in Parana dominion ( Fig. 13 View FIGURE 13 ), separate these two species.
Distribution. From Rio Grande do Sul State, Brazil, and Entre Rios Province, Argentina south to Florida Department, Uruguay; an inhabitant of the Pampean biogeographical province ( Fig. 13 View FIGURE 13 ).
Biological and ecological data. Adults are found in lotic environments such as riffles and deep zones in slow streams surrounded by gallery forest, where adults perch in protected sites, from sea level to 107 meters of elevation. Larvae are shallow burrowers and were found with some adhered detritus (Rodrigues Capítulo 1985).
Remarks. The typification by Rodrigues Capítulo (1985) is misleading and imprecise. In describing C. noval he designated an adult male emerged in the laboratory as holotype without mention of its exuvia (Rodrigues Capítulo 1985, p. 329). Furthermore, the other cited specimens, an adult female and six larvae, were not designated as paratypes, therefore this meets the requirement that “expressly excludes them from the type series” (ICZN 1999, Art. 72.4.6), even though an allotype was mentioned in the abstract. The subsequent citation of the female as allotype by Muzón et al. (2007) should be considered an invalid nomenclatural act. The date of publication of all papers in that volume is stated to be 1984, but the cover states 1985. For this reason, without further investigation, we follow Garrison et al. (2006) considering 1985 as the correct year of publication, while the respective day and month still are pending.
Except by the paler color (“pardo amarillenta”), all characters mentioned by Rodrigues Capítulo (1985, p. 329–332) are not diagnostic of T. noval in accordance with its new status recognized here, and his characters should be considered as examples of individual variation, as they are shared with some T. uncatus . Rodrigues Capítulo (1985) provided no diagnosis, instead comparing a few selected characters with T. uncatus as follows: (1) anterior margin of the postfrons in dorsal view sinuous with a small mesal convexity, while in T. uncatus it is entirely convex; (2) male S9 with a V-shaped mesodorsal invagination vs. lacking invagination in T. uncatus ; (3) spines along posterior carina of S9 extend downward two-thirds of the segment height in the male vs. only over dorsal surface in T. uncatus ; (4) posterior margin of male epiproct almost straight vs. ovoid in T. uncatus ; (5) female distal margin of sternites of S9–10 distinct from T. uncatus . The shape of the anterior margin of the postfrons (character 1) is variable, specimens of T. noval collected from the same site and day showing either a small mesal convexity ( Fig. 5 View FIGURE 5 B) or regularly convex postfrons ( Fig. 5 View FIGURE 5 A) such as illustrated for the holotype of T. uncatus by Rodrigues Capítulo (1985, fig. 1). Additionally, variation in the postfrons in T. uncatus also shows a slightly mesal projection along the anterior margin of postfrons ( Figs 5 View FIGURE 5 C–D). A small mesodorsal invagination in S9 (character 2) is common to all Cyanogomphini including T. uncatus ( Figs 7 View FIGURE 7 B, E, H, K, N). We did not find the shape of invagination to be a diagnostic character. The distribution of spines along S9 (character 3) also is variable among specimens, and in both T. noval and T. uncatus they can extend down two-thirds of the segment height (see Figs 7 View FIGURE 7 J, M). This variation might have lead J. Belle to misidentify some T. noval as T. uncatus because they had the spines extending to the ventral half. Even though in T. noval the distal margin of the epiproct (character 4) appears be straighter than that of T. uncatus ( Figs 7 View FIGURE 7 J, L, M, O, 8G–J), it is not ovoid in T. uncatus , and this structure is similar for both species. Finally, character 5 on female sternites of S9–10 cannot be verified, as differences were not depicted. Although Rodrigues Capítulo (1985) borrowed the holotype of T. uncatus , he adopted the illustrations by Fraser (1947) for his comparisons, possibly due to the poor condition of the holotype (S4–10 lost, see Belle 1980).
The males from municipalities of Guaíba and Pelotas were determined as T. noval , while a couple from Jaguarão municipality, all from Rio Grande do Sul State, Brazil, were determined as T. uncatus by J. Belle in 1992.










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Tibiagomphus noval (Rodrigues Capítulo, 1985)
| Pinto, Ângelo Parise & Almeida, Marcus Vinícius Oliveira De 2016 |
Cyanogomphus noval Rodrigues Capítulo, 1985: 329
| Rodrigues Capitulo 1985: 329 |
Tibiagomphus noval (Rodrigues Capítulo, 1985)
| Rodrigues Capitulo 1985 |