
Listrura costai, Villa-Verde & Lazzarotto & Lima, 2012
|
publication ID |
https://doi.org/ 10.1590/S1679-62252012000300005 |
|
persistent identifier |
https://treatment.plazi.org/id/49053F6D-CB1C-FFCF-FE9A-FE6FFA854A62 |
|
treatment provided by |
Felipe |
|
scientific name |
Listrura costai |
| status |
sp. nov. |
Listrura costai View in CoL , new species
Figs. 1 View Fig and 2 View Fig
Holotype. MNRJ 31917 View Materials , 31.3 mm SL, Brazil, Rio de Janeiro State, Angra dos Reis Municipality, small unnamed stream close to the road RJ-155 (Angra-Lídice), rio Jurumirim basin, 22º55’24.7”S 044º18’51.4”W, 11 Mar 2006, B. B. Costa, C. P. Bove, J. P. Moreira, & W. J. E. M. Costa. GoogleMaps
Paratypes. MNRJ 31535 View Materials , 3 View Materials , 25.9-32.1 mm SL, 2 CS, 20.4-26.5 mm SL, 22 Aug 2005, D. Eberienos, H. Lazzarotto, I. Montenegro & J. Prata . MNRJ 31918 View Materials , 5 View Materials , 14.9 View Materials -29.0 mm SL, 02 Jan 2008, H. Lazzarotto, J. Prata, L. Villa-Verde & S. M. Q. Lima . MNRJ 39620 View Materials , 4 View Materials , 15.0-30.0 mm SL, 27 Jun 2008, D. Almeida, J. Prata & S. M. Q. Lima . UFRJ 7214 , 3 , 28.1-33.5 mm SL , UFRJ 7215 , 4 , 26.0-32.0 mm SL , UFRJ 6577 , 3 CS, 32.0- 34.3 mm SL, 01 Oct 2005, H. Lazzarotto, M. Rheingantz & R. Curcio. All from Brazil, Rio de Janeiro State, Angra dos Reis Municipality, swamp area close to the road RJ-155 (Angra-Lídice), about 1 km from road BR-101, rio Jurumirim basin, 22º55’24.7”S 044º18’37.5”W GoogleMaps .
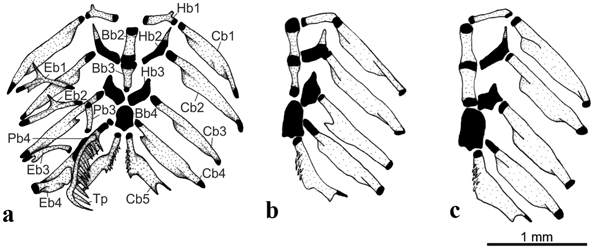
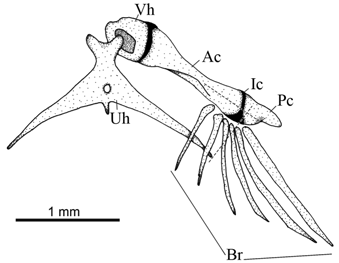
Diagnosis. Listrura costai is autapomorphically distinct from its congeners by possessing a small anterior process on the first hypobranchial (vs. process absent; Fig. 3 View Fig ). The new species is also distinguished from its congeners, except L. nematopteryx and L. picinguabae , by possessing only one pectoral-fin ray (vs. two to four rays). It differs from L. nematopteryx and L. picinguabae by having an inferior row of dots extending only on posterior half of body (vs. over almost the entire body), and posterior tip of ceratobranchial 5 strongly concave (vs. slightly concave or straight; Fig. 3 View Fig ). It further differs from L. nematopteryx by possessing a discontinuous lateral midline row of spots (vs. continuous row), lateral processes of urohyal reaching the anterior region of posterior ceratohyal ( Fig. 4 View Fig ) [vs. reaching the posterior region of posterior ceratohial; see fig. 4B in Villa-Verde & Costa, 2006], more dorsal and anal-fin rays (8-9 vs. 7-8 and 8-9 vs. 6-8, respectively), and more opecular and interopercular odontodes (6-8 vs. 4-6 and 8-12 vs. 5-7, respectively). It further differs from L. picinguabae by possessing a longer head (head length 11.7-15.3% SL vs. 10.7- 11.7% SL), more dorsal-fin rays (8-9 vs. 7), and anal-fin origin at vertical through region from centra of the 31 st to 34 th vertebrae (vs. 34 th to 36 th).
Description. Morphometric data given in Table 1. Body elongate, subcylindrical at anterior portion of trunk, and strongly compressed at caudal peduncle. Dorsal and ventral profiles straight. Skin papillae minute.
Head depressed and trapezoidal. Snout blunt and slightly elongated. Mouth subterminal and narrow. Teeth conical, tips pointed and curved. Two rows of teeth in jaw. Premaxilla teeth 15-26; dentary teeth 13-21. Eyes anteriorly located on head, nearer to snout tip than to opercular patch of odontodes. Nasal, maxillary and rictal barbels well developed. Tip of nasal barbel reaching between posterior margin of interopercular patch of odontodes and posterior margin of opercular patch of odontodes. Tip of maxillary barbel reaching just posterior margin of opercular patch of odontodes. Tip of rictal barbel reaching between middle of interopercular patch of odontodes and posterior margin of opercular patch of odontodes. Anterior nostril opening anteriorly at base of nasal barbel. Posterior nostril located on anterior half of distance between anterior nostril and eye. Interopercular odontodes 8-12, opercular odontodes 6-8; odontodes conical, tips pointed and slightly curved.
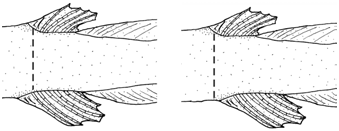
Dorsal and anal fins approximately triangular. Dorsal-fin origin on posterior half of trunk, at vertical between centra of 30 th and 35 th vertebrae. Anal-fin origin anterior to dorsal-fin origin (dorsal-fin origin on vertical through base of 3 rd anal-fin rays) to posterior to it, on vertical through base of 4 th dorsal-fin ray ( Fig. 5 View Fig ), and between centra of 31 st and 34 th vertebrae. Caudal fin elongate with posterior margin rounded, extending dorsally and ventrally to posterior margins of dorsal- and anal-fin bases, respectively. Pectoral fin with single filamentous ray. Pelvic fin and girdle absent. Dorsal-fin rays 8-9, all unbranched; anal-fin rays 8-9, all unbranched; principal caudal-fin rays 11, dorsal-procurrent rays 28-35, ventral-procurrent rays 23-32. Pleural ribs 2-3. Total vertebrae 48-52. Branchial membranes attached only at anteriormost point of isthmus. Branchiostegal rays 6.
Laterosensory system extremely reduced, without supraorbital, infraorbital, mandibular, preopercular, and otic canals ( Fig. 6a). Postotic canal with two pores: anteriormost at vertical through anterior margin of opercular patch of odontodes; posteriormost at vertical just posterior to opercular patch of odontodes. Lateral line on trunk very short, with two pores: anteriormost largest, at vertical just posterior to pectoral-fin base; posteriormost below former in lateral view and just posterior to it.
Osteology. Anterior cornua of mesethmoid straight in dorsal view ( Fig. 6a). Lateral ethmoid with lateral projection. Frontal and parieto-supraoccipital fully joined by sutures; cranial fontanel absent. Sphenotic, prootic and pterosphenoid entirely fused to each other. Vomer without posterior process. Parasphenoid without posterior process. Pterotic with lateral laminar expansion ( Fig. 6a). Basioccipital and exoccipital fully fused to each other and posteriorly fused to Weberian capsule. Co-ossified basioccipital and exoccipital approximately triangular. Weberian capsule with small lateral opening on each side.
Autopalatine nearly square with curved lateral process ( Fig. 6a). Premaxilla triangular. Maxilla elongate.Antorbital short and anteriorly expanded. Supraorbital extremely reduced.
Hyomandibula with elongated and narrow anterior process ( Fig. 6b). Anterodorsal portion of quadrate laminar. Metapterygoid extremely reduced and articulating only with anterodorsal portion of quadrate through cartilaginous block. Preopercle straight. Odontodes disposed vertically on opercle and obliquely on interopercle.
Basibranchial 1 absent ( Fig. 3a View Fig ). Basibranchials 2 and 3 cylindrical, interconnected by cartilage. Cartilages on anteriormost border of basibranchial 2 and posteriormost border of basibranchial 3, respectively. One CS specimen (UFRJ 6577, 32.5 mm SL) with ossified basibranchial element peculiarly modified in crucifix shape ( Fig. 6c; see Discussion). Basibranchial 4 flattened and fully cartilaginous. Hypobranchial 1 cylindrical and with anterior process (see Discussion). Hypobranchial 2 flattened and cartilaginous with anterolateral ossified process. Hypobranchial 3 flattened and fully cartilaginous. Hypobranchial 4 absent. Ceratobranchials 1, 2, 3, and 4 cylindrical, with slight posterior laminar process. Ceratobranchial 5 with strong concavity on posterior tip and dorsally-oriented conical teeth on anterior half. All ceratobranchials with cartilages on tips. Epibranchials 1, 2, and 3 thin; first with anterior and posterior processes, second with small anterior process, and third with small posterior uncinate process. Epibranchial 4 cylindrical with larger posterior laminar process. Epibranchial 5 absent or not evident. Pharyngobranchials 1 and 2 absent. Pharyngobranchial 3 cylindrical. Pharyngobranchial 4 fully cartilaginous and attached to tooth plate. Upper pharyngeal tooth plate with long ventromedially-oriented conical teeth.
Urohyal with narrow pointed and elongated lateral processes reaching to anterior region of posterior ceratohyal ( Fig. 4 View Fig ). Two small condyles on anterior region of urohyal. Deep depression on ventral hypohyal for articulation with urohyal condyle. Anterior ceratohyal cylindrical and constricted at midlengh. Posterior ceratohyal nearly triangular. Interceratohyal cartilage narrow.
Cleithrum flattened, slightly triangular and with large fenestra on central region ( Fig. 6d). Scapulocoracoid narrow, fully cartilaginous and with narrow process on distal portion. Single proximal radial rounded and fully cartilaginous. Single pectoral-fin ray narrow and segmented.
Caudal skeleton compact ( Fig. 6e). Hypurals 1-2 and parhypural fused. Hypurals 1-2 almost completely fused to hypurals 3-5. Uroneural almost completely fused to caudal complex. Preural 1-ural 1 compound sometimes with accentuated dorsal concavity.
Coloration. Body light yellowish gray with minute dark brown to black dots on dorsal portion of flank and dorsum, more concentrated on lateral midline, and laterodorsal portion and dorsal midline of trunk to form three longitudinal rows of small, closely placed dark brown to black spots. Lateral midline spots to form discontinuous row; above lateral midline row of spots, dots often coalesced to form short, dark brown oblique lines near central portion of flank and oblique rows of small dark brown spots on side of caudal peduncle, all following myomeres; below lateral midline row of spots, minute brown dots to form narrow longitudinal row extending to posterior half of trunk. Few minute brown dots below lateral midline of caudal peduncle, usually arranged in oblique rows following myomeres. Venter yellowish white. Head pale brown on dorsal region, light yellowish gray on lateral and ventral regions; dark brown to black chromatophores concentrated over dorsal portion of head and snout, and interopercular and opercular regions, but absent or rare on infraorbital region, making distinctively unpigmented zone between snout, orbit and interopercle; small unpigmented zone on center of dorsal portion of head; dark chromatophores extending to ventral portion of head to produce dark transverse zone on chin and short dark zone on branchiostegal region. Barbels hyaline with brown base. Distal portion of dorsal and anal fins hyaline, proximal portion yellowish hyaline with dark brown chromatophores on fin rays. Narrow marginal black line on fin membrane adjacent to procurrent caudal-fin rays. Caudal fin yellowish hyaline with sparse dark brown chromatophores on fin rays. Pectoral fin hyaline.
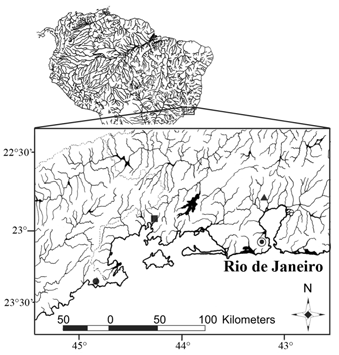
Distribution. Listrura costai is known from two neighboring small unnamed coastal streams adjacent to the road RJ-155, Angra dos Reis Municipality, Rio de Janeiro State, Brazil, both draining into a swampy portion of the rio Jurumirim, close to its mouth ( Fig. 7 View Fig ).
Habitat and ecological notes. Rio Jurumirim is the main canal of a small coastal basin of approximately 65 km 2 which flows through well-preserved areas ofAtlantic forest in the mountain range of the Serra da Bocaina. The rio Jurumirim forms a grassy lowland that becomes partially, and sometimes fully, flooded during periods of high rainfall. In this region, the grass vegetation has replaced the original Atlantic forest probably due to human action.
Most specimens of Listrura costai were collected in a narrow (about 1 m wide) and shallow (about 50 cm deep) clear water stream located in a swampy plain area densely vegetated with grass ( Fig. 8 View Fig ). This site located in rio Jurumirim lowland, just upstream from a paved road (RJ-155), is right next to the pipe that runs underneath the road, draining the stream into a larger tributary of the main river. Most specimens were collected in a shallower portion of this stream (about 10 cm deep) either in the mud bottom or the thin layer of litter covering it ( Fig. 8a View Fig ). Some specimens were also collected at shallowest pools (less than 5 cm deep) adjacent to the stream ( Fig. 8b View Fig ). The locality where the holotype was found has similar characteristics. During the collection of the holotype, the water was clear with pH 6.1 and temperature 25.5ºC at 10:25 am.
Several specimens of L. costai were captured at night or dusk, suggesting a nocturnal habit. Furthermore, two collections during daylight resulted in no specimens captured, opposed to dusk/night time in the same day, when some individuals of L. costai were then found. The new species was found together with Hollandichthys multifasciatus (Eigenmann & Norris) , Gymnotus pantherinus Steindachner , Phalloceros sp. , and Rivulus lazzarotoi Costa , the latter also endemic from rio Jurumirim basin ( Costa, 2007). Other fishes, such as Astyanax taeniatus (Jenyns) , Geophagus brasiliensis (Quoy & Gaimard) , Mimagoniates microlepis (Steindachner) , Mugil sp. , Rhamdia quelen (Quoy & Gaimard) , and Trichomycterus zonatus (Eigenmann) , were also collected in the same stream, however just downstream from the road ( Fig. 8c View Fig ), an area with influence of marine tides, where L. costai was not caught.
Conservation remarks. The extremely restrict distribution, together with a high specificity of its natural habitat (swampy streams in lowland plain areas that rarely occur in southern Rio de Janeiro State watersheds) and relative proximity of human settlements make L. costai prone of becoming critically endangered or even extinct in a very short time period due to effects of human activities or stochastic events. However, no studies about population or geographical range reductions are available. Considering the IUCN categories and criteria to evaluate threatened species (IUCN, 2001, 2010), the actual information about L. costai allows only apply the criterion D2 (area of occupation less than 20 km ²) for the category Vulnerable (VU). Conservation status of L. costai may be better circumscribed with a detailed analysis of its population density. Approval of conservation measures for this area is important not only to preserve this species but also for the conservation of other endemic species, such as Rivulus lazzarotoi .
Etymology. The species epithet is given for Dr. Wilson J. E. M. Costa, in recognition of his significant contributions to Neotropical Ichthyology, including the study with trichomycterid catfishes.
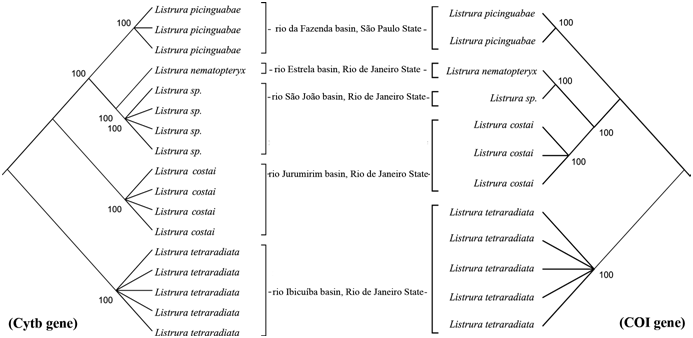
Molecular analyses. Partial sequences of the Cytb gene were taken from 17 specimens which resulted in a matrix with 1085 base pairs (bp) from which 949 sites were conserved, 136 were variable, and 123 parsimony informative. The nucleotide frequencies were 29.8% thymine, 29.6% cytosine, 26.2% adenine, and 14.4% guanine. In COI gene, partial sequences of only 12 specimens were amplified (amplification did not work in one specimen of L. costai , one of L. picinguabae , and three of Listrura sp. ), resulting in a matrix with 789 bp from which 698 sites were conserved, 91 were variable, and 73 parsimony informative. The nucleotide frequencies were 30.0% thymine, 28.3% cytosine, 23.3% adenine, and 18.4% guanine. A single haplotype was found in both mitochondrial markers for each species with more than one sample, except for L. tetraradiata which presented two haplotypes. Values of molecular distances between L. costai , L. nematopteryx , L. picinguabae , Listrura sp. , and L. tetraradiata are given in Table 2. The intraspecific distance values for all species were zero in both mitochondrial genes, except in COI gene of L. tetraradiata that was 0.001 ± 0.001, due to a single nucleotide different on one specimen. Interspecific distance values for Cytb gene range from 0.036 ± 0.006 between Listrura sp. and L. nematopteryx to 0.091 ± 0.009 between L. tetraradiata and L. nematopteryx . For COI gene, the range was from 0.038 ± 0.007 between Listrura sp. and L. nematopteryx to 0.076 ± 0.011 between Listrura sp. and L. tetraradiata . Phylogenetic analyses for Cytb gene obtained 70875 most parsimonious trees, with length of 165 steps, consistency index 0.855263 and retention index 0.957115. For COI gene, 315 most parsimonious trees were obtained, with length of 108 steps, consistency index 0.844444 and retention index 0.936364. Figure 9 View Fig presents two consensus trees obtained from most parsimonious Cytb- and COI-gene trees, respectively, with the bootstrap values for 1,000 pseudoreplicates. Both consensus topologies present a same monophyletic clade ( L. nematopteryx plus Listrura sp. ) well-supported by bootstrap values equal than 100%. Listrura costai also shows well-supported by 100% bootstrap in both topologies.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |