
Rineloricaria quilombola, Chamon & Fichberg, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5194.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:06A9F26D-644A-4F9F-B306-E3C049CF4C57 |
|
DOI |
https://doi.org/10.5281/zenodo.7141894 |
|
persistent identifier |
https://treatment.plazi.org/id/484687A5-2F11-FF96-FF40-2A05FB4593C4 |
|
treatment provided by |
Plazi |
|
scientific name |
Rineloricaria quilombola |
| status |
sp. nov. |
Rineloricaria quilombola , new species
( Figure 1 View FIGURE 1 , Table 1 View TABLE 1 )
Holotype. MZUSP 126901 View Materials , 116.3 View Materials mm SL, Palmas, Taquaruçu, ribeirão Taquaruçuzinho na ponte para o povoado de Taquaruçu Grande, rio Tocantins drainage, -10.31555, -48.21638 16 April 2018, C. Chamon, J. Silva, I. Fichberg and E. Oliveira. GoogleMaps
Paratypes. All from Tocantins River basin. UNT 942 View Materials , 3 View Materials , 87.7 View Materials – 132.6 View Materials mm SL, córrego Imbé , Fazenda Traçadal , Paranã, -12.48333 -48.21666, 21 Mar 1999, Marques, E. et al. GoogleMaps UNT 943 View Materials , 3 View Materials , 71.1 View Materials – 134.3 View Materials mm SL, córrego Cipó , Fazenda Traçadal , Paranã, -12.72805, -48.22944, 26 Jan 1999, Marques, E. et al. GoogleMaps UNT 1094 View Materials , 8 View Materials , 54.3 View Materials –92.0 mm SL, ribeirão Manduca , Porto Nacional , -10.79416, -48.36666, 3 Oct 2001, Marques, E. et al. GoogleMaps UNT 1713 View Materials , 3 View Materials , 44.0–110.3 mm SL, same locality as UNT 943, 4 Aug 2004, Marques, E. et al. GoogleMaps UNT 1723 View Materials , 6 View Materials , 48.4 View Materials –75.0 mm SL, same locality and collect data of UNT 1713 View Materials . GoogleMaps UNT 15542, 2 View Materials , 49.4 View Materials –114.0 mm SL, same locality of the holotype, 26 Jun 2016, Chamon, C., Oliveira, E., Silva, J. and Bezerra, C. GoogleMaps UNT 15711, 6 View Materials , 73.3 View Materials –97.0 mm SL, same locality as holotype, 18 Nov 2016, Chamon, C., Oliveira, E., Pereira , T., Silva, J. and Bezerra, C. GoogleMaps UNT 15794, 2 View Materials , 45.6 View Materials –50.0 mm SL, ribeirão Taquaruçu Grande, na chácara Irecê, Taquaruçu, Palmas, -10.31555, -48.21638, 15 Dec 2016, E. Oliveira and J. Silva. UNT 19246, 1 View Materials , 61.2 View Materials mm SL, collected with the holotype. GoogleMaps UNT 19262, 3 View Materials , 59.6 View Materials – 111.5 View Materials mm SL, same locality of the holotype, 26 Apr 2016, Chamon, C., Oliveira, E. and Silva, J. GoogleMaps UNT (uncatalogued), 4, 50.1–62.3 mm SL, ribeirão São João, Porto Nacional, -10.73660, -48.29243, 23 May 2022, Pelicice, F. Rocha, T., Fontoura, F., Parente, A.L. and Chamon, C GoogleMaps .
Non-types. UNT 945 View Materials , 1 View Materials , 98.6 View Materials mm SL, same locality of UNT 942 View Materials , 26 Sep 2001, Marques, E. et al . UNT 956 View Materials , 1 View Materials , 107.6 View Materials mm SL, same locality as UNT 942, 25 Aug 1998, Marques, E. et al . UNT 1029 View Materials , 20 View Materials , 53.9 View Materials – 89.4 View Materials mm SL, córrego Taboca , Paranã, -12.5927778000, -48.1036111000, 7 Aug 2000, Marques, E. et al GoogleMaps . UNT 1038 View Materials , 1 View Materials , 72.9 View Materials mm SL, same locality as UNT 945, 3 Oct 2001, Marques, E. et al GoogleMaps . UNT 7190 View Materials , same locality as UNT 942, 15 Aug 2005, Marques, E. et al GoogleMaps . UNT 7911 View Materials , 10 View Materials , 40.0–77.6 mm SL, same locality of GoogleMaps UNT 943 View Materials , 24 Aug 2007, Marques, E. et al .
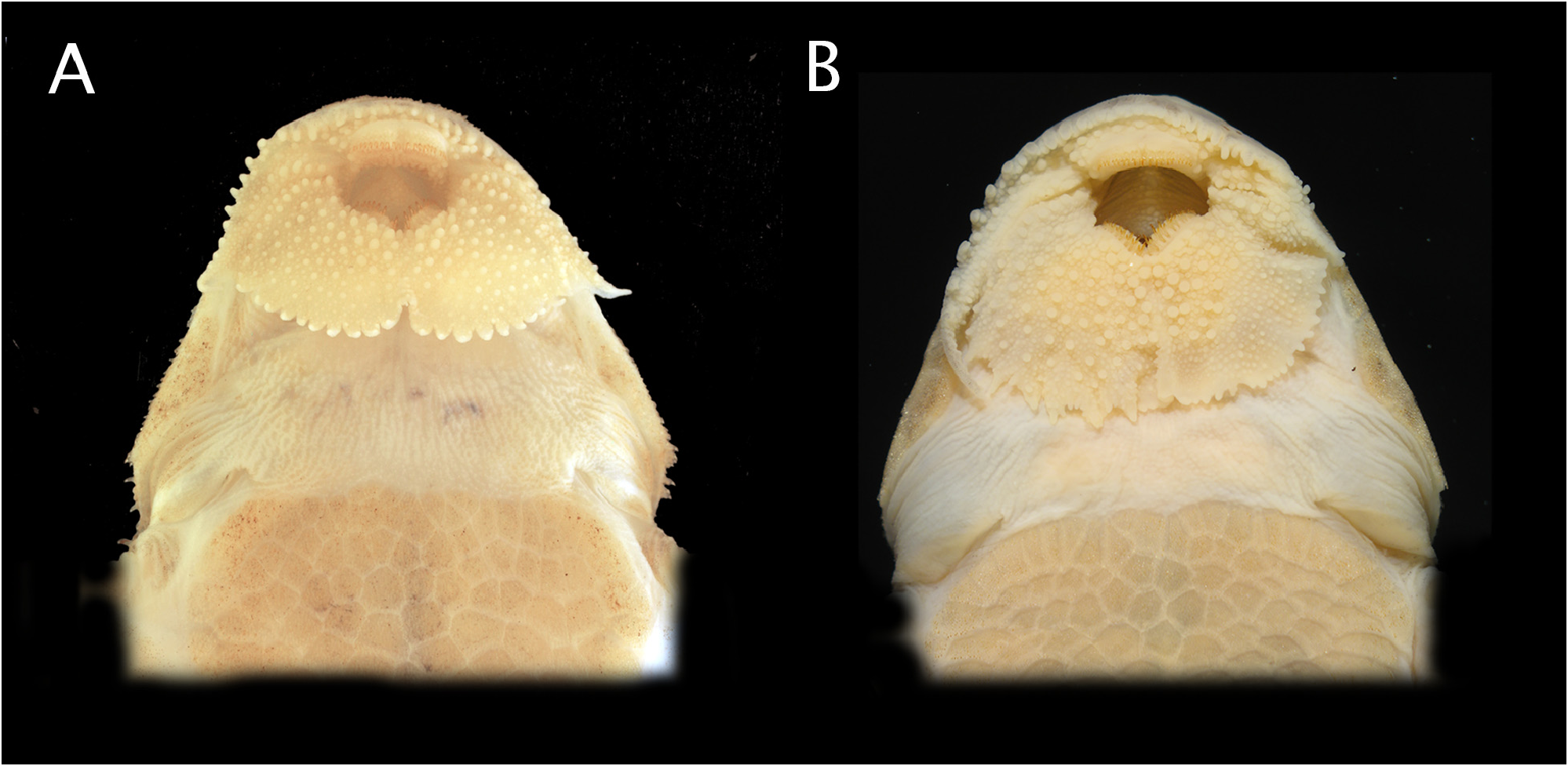
Diagnosis. Rineloricaria quilombola is diagnosed among its congeners by a greater number of premaxillary teeth (up to 15 vs. up to 10 in all congeners, except R. aequalicuspis and R. castroi with 15; R. osvaldoi with 13 and R. uracantha with 12); and dentary teeth (up to 15 vs. up to 10 in all congeners, except R. aequalicuspis with 20, R. osvaldoi with 13; R. castroi and R. uracantha with 12). From R. aequalicuspis , it can be differentiated by having teeth with a principal inner cusp and an accessory smaller one (vs. teeth with inner and outer cusps of approximately the same size). From R. castroi , it is distinguished by color pattern without dark stripes on the pectoral, pelvic and anal fins (vs. pectoral, pelvic and anal fins with conspicuous dark stripes). From R. hasemani it is distinguished, by having the edge of snout straight in lateral view, absence of filaments on lower lip and by color pattern lacking a dark stripe on dorsal fin (vs. edge of snout upturned in lateral view, absence of filaments on lower lip and dark stripe on dorsal fin present). From R. lanceolata , by a wider body at cleithral width 16.5–20.1% of SL (vs. 13.8–16% of SL); presence of five lateral plates series in longitudinal rows below dorsal fin (vs. four lateral plates series); base of pectoral and pelvic-fins completely ochre or light brown without a hyaline area (vs. base of pectoral and pelvicfins with a hyaline circular area); and by the absence of a dark stripe in all fins, running parallel to the first rays (vs. dark stripe present). From R. osvaldoi , the new species is further diagnosed by a lower head depth (30.8–40.5 % vs. 42.2–61.3 % of HL), by having shorter snout length (45.5–51.4 vs. 50.3–68.8% of HL); triangular head in dorsal view (vs. rounded head in dorsal view); lower lip with elongated and organised rounded-shape fringes (vs. lower lip with short and unorganised triangular fringes); lower lip short not covering the gular area (vs. lower lip elongated, covering about 50% of the gular area); upper lip margin with long, rugged and regular papillae (vs. upper lip margin with short and inconspicuous papillae) ( Fig. 2 View FIGURE 2 ); head with regular rows of odontodes (vs. odontodes not forming regular rows); dimorphic male lateral head with short and spaced apart odontodes (vs large and very close hypertrophied odontodes).
Description. Morphometric data in Table 1 View TABLE 1 . Head and body strongly depressed. Dorsal profile of head triangular with tip of snout and distal margin of opercle rounded. Snout tip straight, not raised in lateral view. Snout with very small and elliptical naked area short and not reaching the first pore of infraorbital ramus of sensorial canal. Dorsal profile concave from tip of snout to dorsal-fin origin, and straight from this point to caudal-fin origin. Ventral profile straight from tip of snout tip to caudal-fin origin. Greatest body depth at the posterior border of supraocciptal; lowest body depth at caudal peduncle.
Head short and very depressed. Odontodes of head small to moderate size, forming conspicuous ridges between nostrils on posterior nasal plates to posterior margin of parieto-supraoccipital and compound pterotic.Six infraorbital, infraorbital 1 with sensorial pore exposed ventrally, infraorbitals 2, 3 and 4 raised convexly from anterior region of snout to eye. Predorsal plates and first three lateral plates of dorsal series slightly keeled, covered with small odontodes. Eyes elliptical with a large and deep postorbital notch, slightly larger than half of the horizontal diameter of the orbit.
Mouth opening large. Upper lip short and separated from the naked area of snout by an extremely thin row of plates covered by tiny odontodes; margin of upper lip adorned with long, rugged and regular papillae. Two rows of papillae between anteroventral border of upper lips and anterior border of premaxillary-ramus; lower lips covered by irregular sized papillae unorganised and regularly distributed around oral cavity; edge of lower lips with elongated and organised triangle-shape fringes. Maxillary barbel moderate in size, reaching the lower lip distal margin. Barbel adorned with very small papillae. Teeth acute and strongly bicuspidate; dentary teeth slightly larger than premaxillary; premaxilla with 8(8), 9(5), 10(3), 11(4)*, 12(1), 13(3) or 15(1) teeth and dentary with 8(6), 9(1), 10*(4), 11(7), 12(4), 13(1), 14(1) or 15(1) teeth; cuspids orange coloured, accessory cuspid almost the same size of principal one.
Body covered by 27(7) or 28*(18) plates on median series, coalescence on 15(6), 16(9)* or 17 (10) and, 8(9), 9(9)* or 10(7) lateral abdominal plates. Mid-dorsal series with 2(15)*, 3(5) or 4(5) plates. Lateral line complete. Five longitudinal rows of plates at dorsal-fin origin. Lateral plates weakly keeled with odontodes along the lateral line pores slightly more developed than those at the rest of the body. Ventral region totally covered by plates from cleithrum to caudal peduncle. Ventral plates well organised in three sections. Anterior section with small, quadrangular plates on the pectoral girdle area. Second section includes large and trapezoidal plates between pectoral and pelvic girdles. Third section represented by the preanal shield formed by three large plates surrounding polygonal preanal plate.
Dorsal fin (ii,7), dorsal-fin spinelet present, locking mechanism not functional. Dorsal-fin base with four to five plates. Pectoral fin (i,6), adpressed unbranched ray slightly surpassing pelvic-fin origin. Pelvic fin (i,5), depressed unbranched ray reaching anal-fin origin. Anal fin (i,5), with two plates on base. Caudal fin (i,10,i) emarginated, with short and thin filament on the upper caudal-fin ray. Lower caudal-fin ray filament absent.
Coloration in alcohol ( Fig. 3 View FIGURE 3 ). Body background with light brown to beige coloration. Dorsal surface with five to six transverse dark brown bands: first one located at second or third dorsal-fin ray, extending toward pectoral-fin origin; second one at the distal edge of dorsal fin when adducted, extending toward to pelvic-fin origin; third one located at the distal edge of the pelvic fin when adducted; fourth, fifth and sixth ones located at caudal peduncle. First and second longitudinal bands occasionally faint, posterior ones darker and more conspicuous ( Figs. 1 View FIGURE 1 and 3 View FIGURE 3 ). Second band occasionally wider in some specimens ( Fig. 3A View FIGURE 3 ). Presence of sixth band on the caudal peduncle observed only in few specimens under 63 mm of SL ( Fig. 3C View FIGURE 3 ). Dorsal surface of snout and head covered with irregular dark brown dots; more concentrated at nasal, frontal and posterior margin of the parieto-supraoccipital and compound pterotic. Posterior margin of compound pterotic with dark-brown to black tiny spots around the first pores of lateral-line canal. Specimens with less than 70 mm of SL present conspicuous dark brown lines between nostrils (along the nasal bones) and dark brown lines behind the postorbital notch and extending to supraoccipital ( Fig. 3C View FIGURE 3 ). Lines less conspicuous or completely absent in larger specimens. Pores of sensorial canal of head and lateral line highlighted by dark brown or black chromatophores. Ventral surface background with beige coloration without dots, spots or bands ( Fig. 1C View FIGURE 1 ). Dorsal surface of upper lip with two longitudinal light brown stripes, lower lip beige without dots or blotches. All fins with light brown or beige coloration covered with dark brown dots that usually run in parallel to fin-rays, dark spots could be faint on pelvic and anal fins; caudal fin with a conspicuous dark brown blotch on base and faint darker band on distal edge ( Figs. 1 View FIGURE 1 and 3 View FIGURE 3 ).
Coloration in life. Live coloration is very similar to color in alcohol, with a dorsal background ranging from ochre to grey and faint dorsal dark brown transversal bands. Dots and lines on snout and head are more conspicuous, as well as the dark brown dots on fins surface ( Fig. 4 View FIGURE 4 ).
Sexual dimorphism. Head margin of mature males triangular, with short and spaced apart hypertrophied odontodes extending from postrostral plates to opercle, along margin of head; unbranched pectoral-fin ray and first four branched rays weakly covered dorsally by well-developed odontodes ( Fig. 5 View FIGURE 5 ).
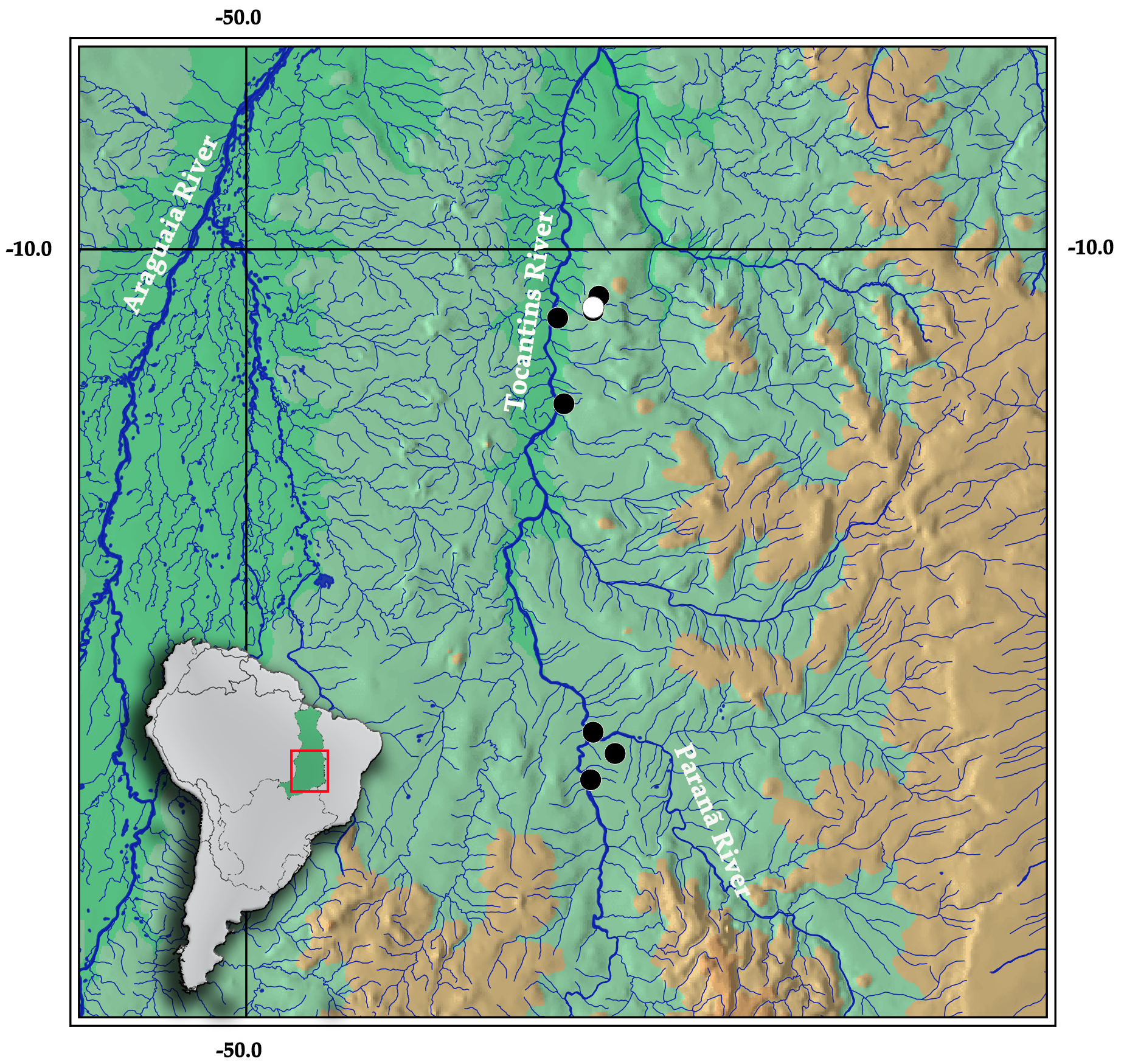
Distribution. The new species is known from upper and middle stretches of Tocantins River basin, in the municipalities of São Salvador, Paranã, Palmas (at Taquaruçu district) and Porto Nacional, Tocantins State, Brazil ( Fig. 6 View FIGURE 6 ).
Ecological and conservation notes. Specimens from Taquaruçu were collected in streams (Taquaruçuzinho and Taquaruçu Grande, middle Tocantins River) with moderate to strong currents with clear water and background with rocks, stones, gravel and sand ( Fig. 7A View FIGURE 7 ). Although the Palmas region is an area of influence of the Lajeado reservoir, at Taquaruçu district, the area is relatively preserved since it is sheltered within the environment protected area (APA) of Serra do Lajeado. Thus, these streams have preserved riparian forest in most of their courses. Still, there is concern due to water withdrawal of the headwaters for agricultural irrigation and recreation activities.
Other localities where the species is distributed, include Cipó, Taboca and Imbé streams, that are located at the influence area of São Salvador and Peixe Angical reservoirs (upper Tocantins River), near the confluence of Paranã and Tocantins rivers; and Manduca and São João streams ( Fig. 7B View FIGURE 7 ), located upstream Lajeado reservoir, at Porto Nacional municipality. Most of nearby areas of these streams were strongly altered by the implementation of reservoirs (e.g. São Salvador, Peixe Angical and Lajeado), and agricultural expansion. Even so, the streams are preserved with moderate riparian forest on its courses
According to GeoCAT analysis, the new species Extent of Occurrence (EOO) is 4,822 km 2 and Area of Occupancy (AOO) is 28.000 km 2, based solely on these criteria, IUCN criteria suggest that the species is Endangered (EN). However, there was no capture effort since 2007, in addition, other biological aspects of the new species, such as population size and generational time are not known. Thus, we suggest that Rineloricaria quilombola should be categorised as deficient data (DD) according to the International Union for Conservation of Nature categories and criteria ( IUCN, 2021).
Etymology. The specific epithet (a noun in apposition) is in honour of all the remaining Quilombola population of the state of Tocantins, which hosts more than 40 communities called Quilombos. Quilombos were specifically communities created by fugitive enslaved African-Brazilian people in Brazil. These places became centers of resistance for enslaved people and their descendents. These communities are legally protected by the Brazilian government yet they are under constant threat related to the expansion of agribusiness and land speculation. Part of the distribution area of the new species is inserted or close to some of these communities.





| UNT |
Universidad nacional de Tucumn |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |