
Grammomys selousi, Denys & Lalis & Lecompte & Cornette & Moulin & Makundi & Machang & Volobouev & Aniskine, 2011
|
publication ID |
https://doi.org/ 10.5252/z2011n1a1 |
|
persistent identifier |
https://treatment.plazi.org/id/475DBC14-FFBD-6624-FF4B-08B3EB88F936 |
|
treatment provided by |
Felipe |
|
scientific name |
Grammomys selousi |
| status |
sp. nov. |
Grammomys selousi n. sp.
( Figs 9-16 View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG )
TYPE MATERIAL. — Tanzania. Holotype: Kingu Pira, Kichi Forest , line F, 8.VII.2003, 1 adult Š KP171 ( MNHN-CG2007-1231 ).
Paratype: same site as holotype, 9.VII.2003, 1 young Š KP186 ( MNHN-CG 2007-1252).
Both specimens have been prepared as skin, skull, liver in ethanol, fibroblast cultures. External dimensions are summarized in Table 10, skull dimensions in Table 11. Standard karyotype is displayed in Figure 11. View FIG
TYPE LOCALITY. — Tanzania, Lindi Region, Ngarembe village, Kichi Forest (08°27’14”S, 38°39’10”E).
DIAGNOSIS. — Medium sized Grammomys (HB: 79- 107 mm) with tail length representing 147 and 156% of HB, HF: 23-23.5 mm. It is characterized by a yellow-light brown dorsal short pelage and a cream-white ventral one. The tail is yellow and does not display a terminal tuft but has some pale scattered longer hairs (2-3 mm long) at the extremity. There is no subauricular tuft visible. The skull length is 24.88-29.65 mm, nasal length relatively short (7.84-11.09 mm) and braincase round and wide delimitated by rectilinear V-shaped fronto-parietal crests. There is a moderate development of stephanodonty in the molars. On the upper M1, t1 linked to t2, presence of a stephanodont crest with t7 not visible on the t4. The t9 and t8 are round and related by a little oblique crest and there is a well-developped posterior cingulum. On the labial side of the lower molars there is a continuous cingular margin with small cusplets development. The upper M3 is small and round with well marked T1. It is characterized by the following chromosome formula: 2n = 49-50, FNa = 56.
At equivalent age teeth wear classes 3 to 5, it is different from G. dolichurus , G. caniceps , G. macmillani in being larger (skull and external dimensions). It is smaller than G. cometes , G. poensis , G. gigas , G. dryas , G. ibeanus , G. brevirostris . It has about the same body and tail length proportions as G. aridulus but with a larger ear and HF. It is about the same size as G. butingi and G. minnae type specimens. It differs from G. minnae , G. caniceps , G. dolichurus s.l. and all other karyotyped specimens by its chromosome formula. G. ibeanus is larger, has a brown dull pelage with long hairs, the tail tuft is more visible and at the last third of the tail one finds long brown hairs (4-5 mm long). Grammomys ibeanus displays a longer nasal and less globular braincase; its frontoparietal crests are not rectilinear. Grammomys brevirostris has an isolated t1 on the upper M1 and a longer rostrum and masseteric plate.
DESCRIPTION
Yellowish-brown pelage on the back with grey bases (about 9-10 mm over middle rump). The yellowbrown colour is less intense on the flanks, which are lighter. There is a marked demarcation line with the nearly pure white belly. The upper part of the head has the same colour as the back, and the mouth region is white like the belly. Tail notably longer than head and body (147-156%), yellow to light-brown in colour terminated at the tip by longer hairs of the same colour (2-3 mm) ( Fig. 9 View FIG ). There are no hairs on the tail at the base, small hairs appear about the middle of the tail and increase progressively in length till its end but they remain scarce.Top of fore and hind feet of the same colour pale yellow white turning to pale brown on the underside, ungual tufts white. Hind feet short but wide. Digit 5 longer than 1, plantar surfaces naked, with 6 small cushion pads on the hindfoot.
The skull shows the Grammomys microdont tooth pattern ( Fig. 10 View FIG ). The braincase is rounded and inflated in the parietal region, the interorbital region is marked but not very narrow, the rostrum is quite short and narrow. There are supra-orbital ridges starting in the interorbital constriction region on the border of the orbital foramen at the level of the naso-frontal suture and ending in the middle of the parietal bones just above the end of the zygomatic arch.In lateral view, the braincase is convex especially in the parietal region and the occipital region where it makes a re-entrant angle with the inter-parietal plan. The tympanic bullae are not inflated quite proportional to the small size of the skull. There are long and narrow incisive foramen ending at the anterior cingulum level of the first molar. The very small posterior palatal foramina are situated at the level of the first lobe of second molar. The mesopterygoid fossa is narrow and long, its anterior border rounded. The pterygoid fossa is shallow with no obvious ventral part of foramen ovale visible. The pterygoid bridge is low and the foramen ovale small. There is a small round opening of the alisphenoid canal.The middle lacerate foramen is small and narrow; the carotid canal is long and thin (more than 2 mm). In lateral view the mastoid part of the tympanic bullae is slightly inflated. The temporal ridges are well marked and vertical. The mandible is short.
Incisors are yellow without any striation and opistodont. The molars display moderately developed stephanodont crests on the t1 and t3. The t4 cusp of the upper M12/ is prolongated by a crest not related to the t8 but there is no visible t7 on it. On the upper M12/ the t8 is high and a posterior cingulum crest is visible, the t9 is small and very round and situated very anteriorly, close to the t6 and there is a little oblique crest relating the t9 to the t8. From the t8 reaching the labial border of the M12/ one can see a posterior cingulum crest. The lower molars display a continuous cingular crest on the labial margin of the lower M/1. The Cp is large on the M/12. Some stephanodont crests are seen on the second lobe of the M/12 and the prelobe of the lower M/1 harbours a well-developed tma. The M3/3 are relatively small and have a round aspect ( Fig. 11 View FIG ). The upper M3/ displays a t1 and a t3. There is no antero-labial cingulum on the lower M/3.
The adult specimen of the new species is smaller than G. gigas , G. dryas and G. cometes , G. kempi , G. ibeanus from Kenya, Sudan and RSA ( Tables 10, 11). The adult specimen fits within size range of G. dolichurus , G. macmillani from Tanzania and Sudan but has a longer hindfoot. It has broadly the same size than G. aridulus which is a very old specimen (dental wear stage 6). It also fits within the size variation of G. dolichurus from Natal in RSA. But, a comparison with the type specimen of G. dolichurus from BMNH shows that our specimen has a more globular skull and a very short nasal ( Tables 10, 11). The hindfoot length of G. selousi n. sp. enters into the variability of G.dolichurus , G. aridulus and G. minnae , as assessed by Hutterer & Dieterlen (1984), but is notably smaller compared to the types of G. poensis , G. gigas and G. ibeanus . The holotypes of G. cometes and G. gigas , are old adults characterized by advanced wear stages of the molars but they display much larger upper molar rows than the new Kingu Pira species. The type specimens of G.macmillani , G.surdaster and G.dryas have much smaller molars and the type specimens of G. surdaster display the smallest M3/3. The last three species are also characterized by a relatively moderate stephanodonty. The new Selous Grammomys also differs from G. brevirostris by its larger skull, longer rostrum, and like other east and south African species by the t1 not isolated from t2.
External and skull quantitative comparisons
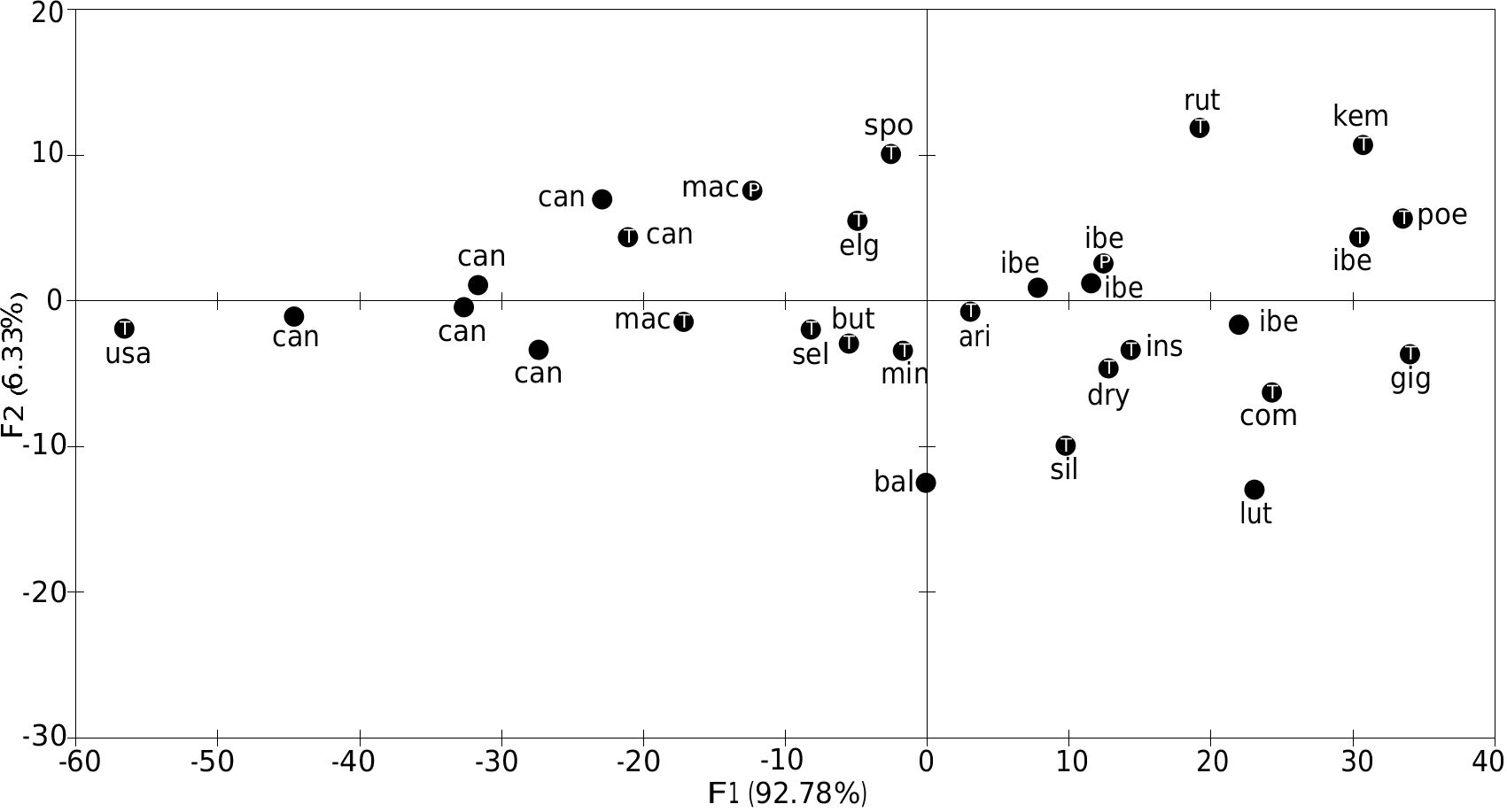
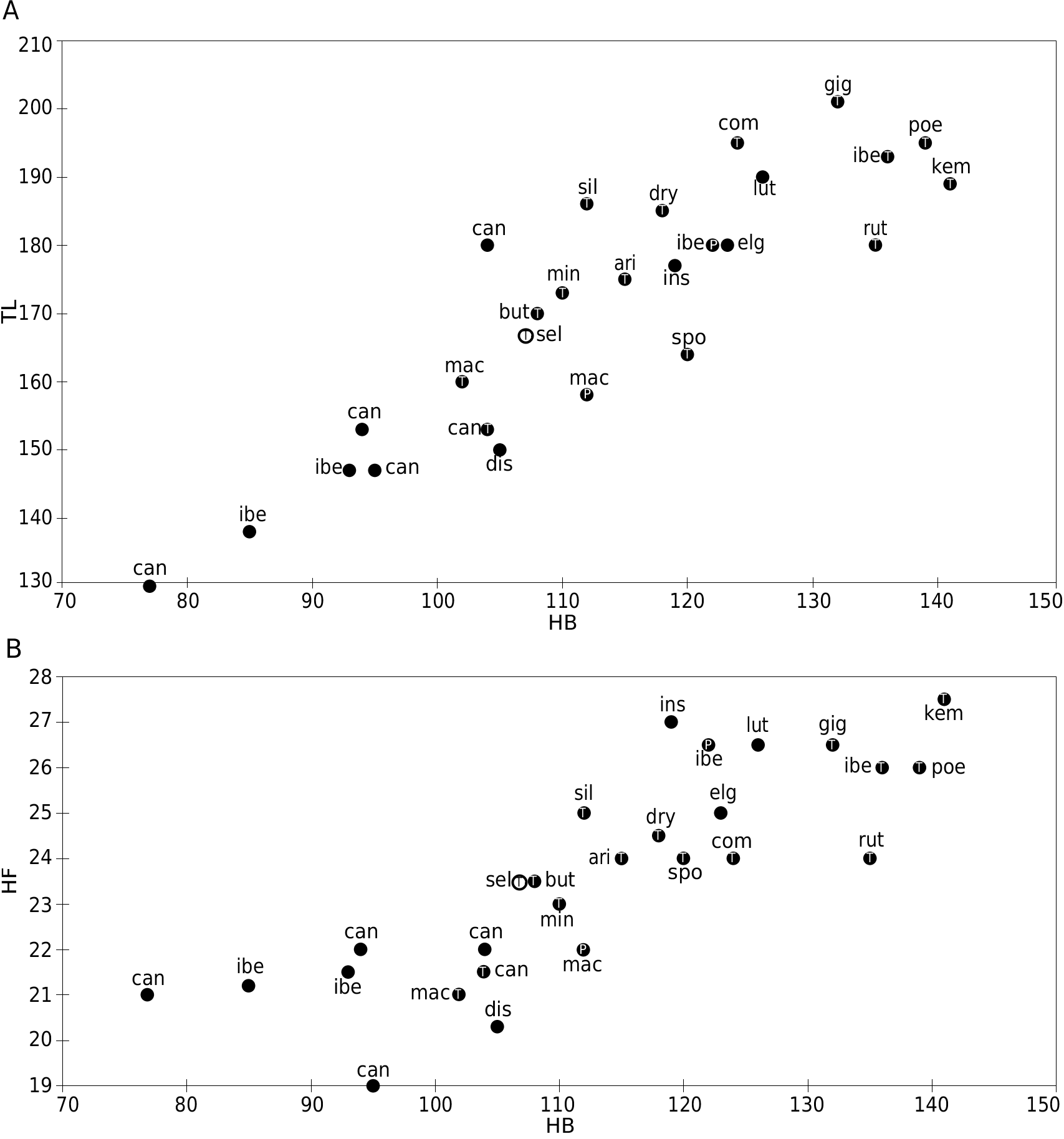
The PCA performed upon the four standard external measurements for all the holotypes available (after Table 10) displays, on the graph of axes 1 and 2, 99.11% of the variability. Axis 1 is positively correlated with all the variables and represents a size axis. Thus the PCA graph shows the relative intermediate size of G. selousi n. sp. ( Fig. 12 View FIG ), which is larger than G. caniceps and G. macmillani and smaller than G. poensis , G. ibeanus , G. gigas , G. dryas , Grammomys kempi , G. cometes and G. rutilans holotypes. G. selousi n. sp. is here close to G. aridulus , G. macmillani , G. minnae and G. butingi . Compared to the latter holotypes, G. selousi n. sp. is at wear stage 4, while G. aridulus and G. butingi display very worn molars (stage 6), G. macmillani holotype is at wear stage 3 and the G. minnae one is at wear stage 4. The scatter plots display the same pattern and show the proximity of G. selousi n. sp. with G. butingi ( Fig. 13 View FIG ).
We performed also a PCA on 11 skull measurements based upon 50 individuals only (after data in Table 11). The graph of axis 1 and 2 displays 89.51% of the total variability. Axis 1 is explained and correlated positively with LGT, LMDB, LNAS, HMDB, WZYG while axis 2 is explained by WBR and LS13, LI13. We observe here the relatively wide variability of G. dolichurus from RSA while G. selousi n. sp. is again in the middle of the graph and close to the type specimen of G. aridulus . The holotypes of G. gigas , G. cometes and G. poensis (the largest) are opposed along axis 1 to the smallest G. caniceps ( Fig. 14 View FIG ). In order to include some of the damaged holotypes to the analyses, we used also bivariate analyses and the scatter plot of LGT versus WZYG confirms the large size of G. ibeanus , the small size of G. macmillani as well as the very wide skull of G. brevirostris ( Fig. 15A View FIG ). By looking at the molar row length versus skull length we confirm the same result and do not see any microdonty trend (reduction of LS13 length) among the species we compared ( Fig. 15B View FIG ).
Karyological description
The new species is characterized by its peculiar chromosome formula and morphology.Two specimens of our study possess 49and 50chromosomes respectively and similar FNa = 56 ( Fig. 16 View FIG ). Th e variability of diploid numbers results from Robertsonian polymorphism well-known in mammals and especially in rodents and previously described for Somalian Grammomys ( Roche et al. 1984) . Although karyotypically the Kingu Pira individuals display some affinities with the specimens called G. kuru (ex rutilans ) by Matthey (1963) from Pointe Noire (CDR), they are clearly different from all earlier descriptions of karyotypes ( Table 12) in Grammomys . Notably, this karyotype is strongly different from those of G. dolichurus from RSA, G. butingi and G. poensis from Ivory Coast, G. gazellae from CAR, G. minnae of Ethiopia and from that of the Grammomys sp.described in northern Tanzania by Fadda et al. (2001). There still is some debate in Grammomys taxonomy ( Corti et al. 2005). According to Musser & Carleton (2005), G. poensis (ex rutilans ), whose type specimen comes from Bioko island ( Equatorial Guinea) and was found in Ivory Coast by Tranier & Dosso (1979), is characterized by 2n = 36 chromosomes.This chromosome formula is different from that of the Congo specimen from Pointe Noire, which would better be attributed to G. macmillani sensu Musser & Carleton (2005) . Therefore, the G. poensis (ex rutilans ) sensu Musser & Carleton (2005) is probably a complex of sibling species whose revision is urgently needed. When looking at all the karyotypes formulae, we see that sibling species may also exist among G. dolichurus , G. surdaster and G. butingi complexes.
There is no available karyotype for G.aridulus which comes from Darfur region ( Sudan) but the latter is greyish brown in colour and proportionaly has a long tail (152%). Because the holotype is at wear stage 6, we infer that the repeated proximity with G. selousi n. sp., which is at stage 4, is not reflecting the mean size of G. aridulus populations which must be smaller than G. selousi n. sp. Moreover, there is a considerable geographic distance between Kulmé and Kingu Pira (about 3000 km) and both places are respectively situated in different ecoregions (sudanian savanna for G. aridulus versus Miombo woodland for G. selousi n. sp.).Similarly, G. brevirostris is not known by karyotypes but its wider skull and shorter rostrum indicate that it is very different from G. selousi n. sp.
In conclusion, because our new specimens display important morphological and size differences with holotypes specimens and chromosomal ones of other Tanzanian forms, we propose to attribute our Selous individuals to a new Grammomys species pending further molecular and cytogenetical analyses in the genus.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.