
Hymenocephalus sp. 2
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3888.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:1B437AE1-CF28-4C1B-95B6-C31A295905A0 |
|
persistent identifier |
https://treatment.plazi.org/id/463A8F36-FFF1-FFBE-1297-9E0EE183FDEF |
|
treatment provided by |
Felipe |
|
scientific name |
Hymenocephalus sp. 2 |
| status |
|
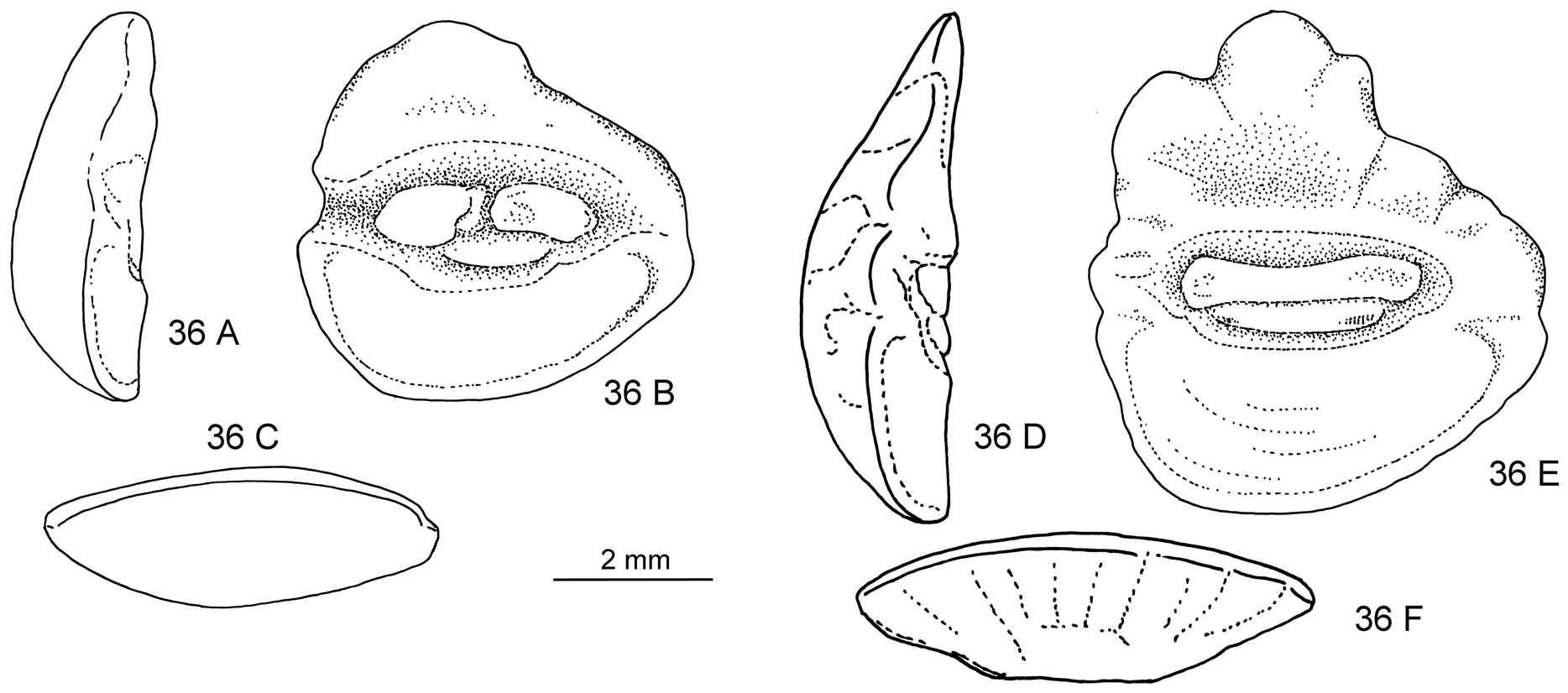
Figs. 36D–F View FIGURE 36 , 37 View FIGURE 37
Material examined. 1 specimen USNM 149049 About USNM , 05°57’S, 81°50’E, 0– 732 m GoogleMaps .
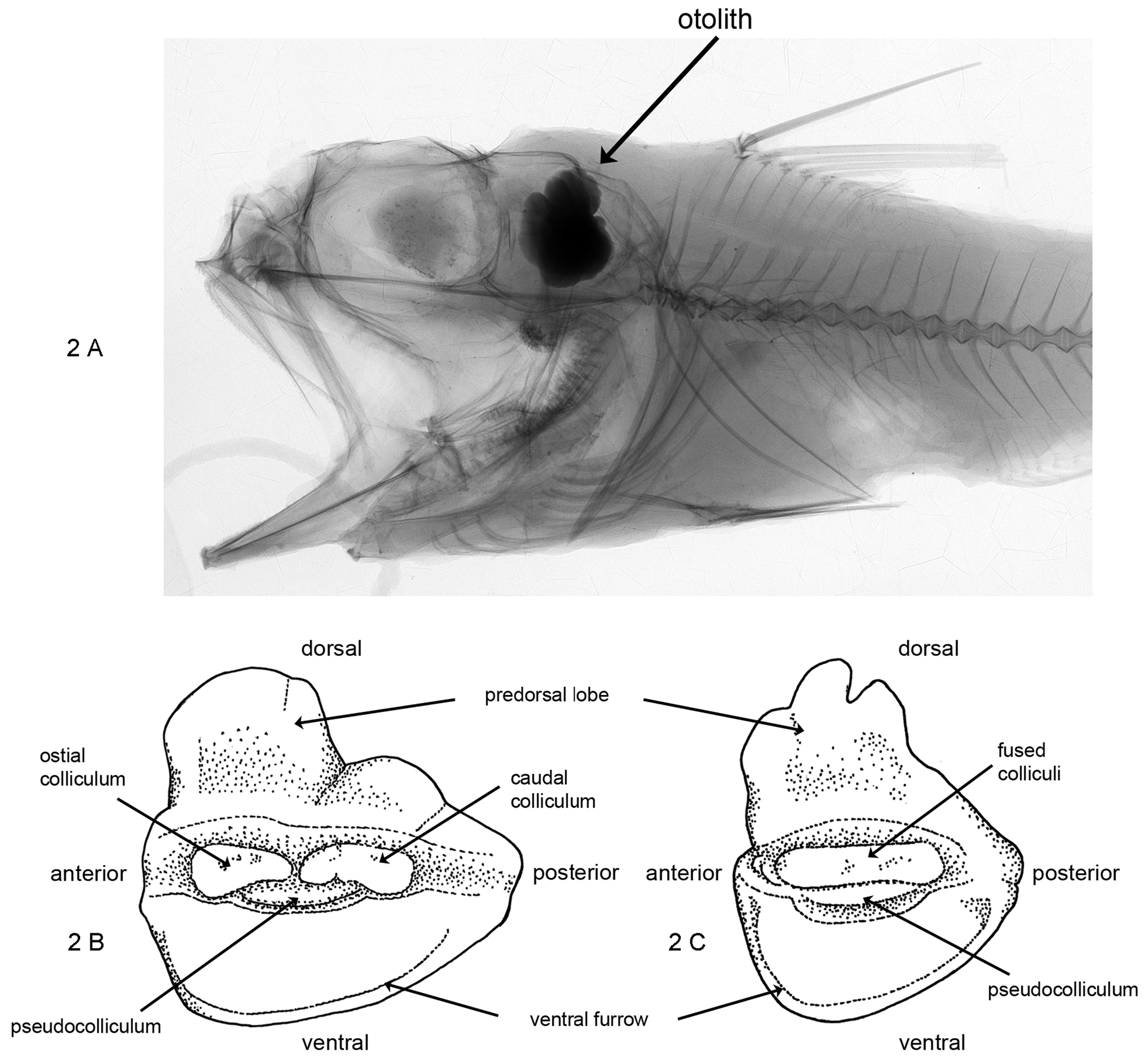
Description. Otolith large; OL:OH = 0.9; OH:OT = 3.5. Dorsal rim with large, moderately high, broad, strongly undulating predorsal lobe, distally marked by small indentation; posterior tip rounded, positioned at about level of sulcus termination; ventral rim deep, regularly curved, smooth, deepest anterior of the middle; anterior rim high, nearly vertical. Inner face slightly convex, with median sulcus. Colliculi completely fused, narrow, terminating far from anterior and posterior tips of otolith; pseudocolliculum very long. TCL:PCL = 1.35. Dorsal depression moderately large, indistinct; ventral furrow moderately distinct, close to ventral rim.
Discussion. This otolith is from the only specimen of the genus collected off the western coast of the Americas (as noted in Cohen et al., 1990). The relatively large fish is almost completely disintegrated, but contained a well-preserved otolith, which closely resembles those of the striatissimus Group, for instance H. striatissimus or H. torvus . The fish either represents a stray specimen of one of the two species, which cannot be identified from the otolith alone, or an unrecorded species.
Fossil record
Despite their Recent abundance, the fossil record of the genus Hymenocephalus is almost non-existent. The only valid record to date is the otolith-based species H. rosenkrantzi Schwarzhans, 2003 from the Middle Paleocene (62–59 Ma) of Denmark. This record confirms a geologically rather early occurrence of the genus together with a few other macrourid species of the same age. Hymenocephalus rosenkrantzi is characterized by a rather elongate shape (OL:OH = 1.5–1.65), a moderately developed predorsal lobe, the caudal colliculum being slightly longer than the ostial colliculum and a short pseudocolliculum positioned only inside the collum and with the ventral margin of the collum being slightly indented. This clearly is plesiomorphic otolith morphology, probably positioned phylogenetically near the dichotomy of the genera Hymenogadus and Hymenocephalus .
Phylogenetic interpretation
Geographical distribution and speciation ( Figs. 5 View FIGURE 5 , 7 View FIGURE 7 , 12 View FIGURE 12 , 16 View FIGURE 16 , 21 View FIGURE 21 , 27 View FIGURE 27 , 29 View FIGURE 29 , 37 View FIGURE 37 )
Fishes of the genera Hymenocephalus , Hymenogadus and Spicomacrurus occur worldwide in tropical to temperate seas except along the continental margin of the eastern Pacific (one specimen found off Peru; see Hymenocephalus sp. 2 above). They live benthopelagically at depths chiefly between 300 and 1000 m over the continental slopes, oceanic island slopes, and above seamounts and guyots. They appear to be largely missing from open oceanic midwater environments.
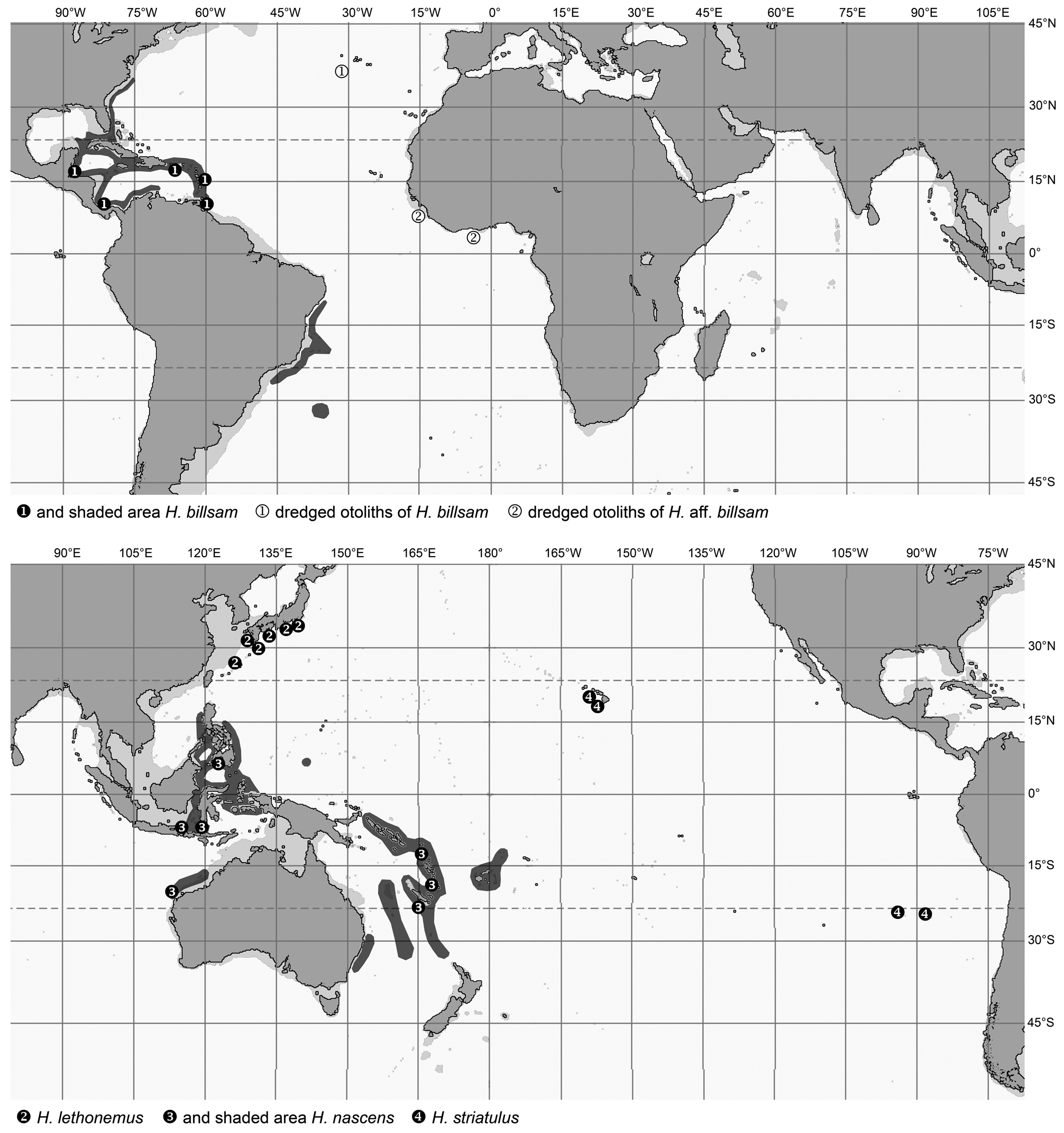
Such a mode of living might support certain trends of geographic regionalization observed in the various species of the genera, although other species show a very wide, nearly cosmopolitan tropical distribution without any recognizable taxonomic diversification. The most outstanding ones in respect to wide geographical distribution are Hymenogadus gracilis and Hymenocephalus aterrimus . The distribution pattern of both species is discontinuous, patchy, but interestingly covers a certain proportion of common terrain in the Caribbean, the southwestern Pacific and the central Pacific Islands of Hawaii and Sala y Gomez seamounts, despite the fact that H. gracilis generally occurs in shallower water than H. aterrimus . In fact, H. gracilis is present over the Sala y Gomez Ridge, but replaced in Hawaii by the only other congener, Hymenogadus tenuis , while H. aterrimus is present off Hawaii, but replaced by the closely related H. sazonovi over the Sala y Gomez Ridge.
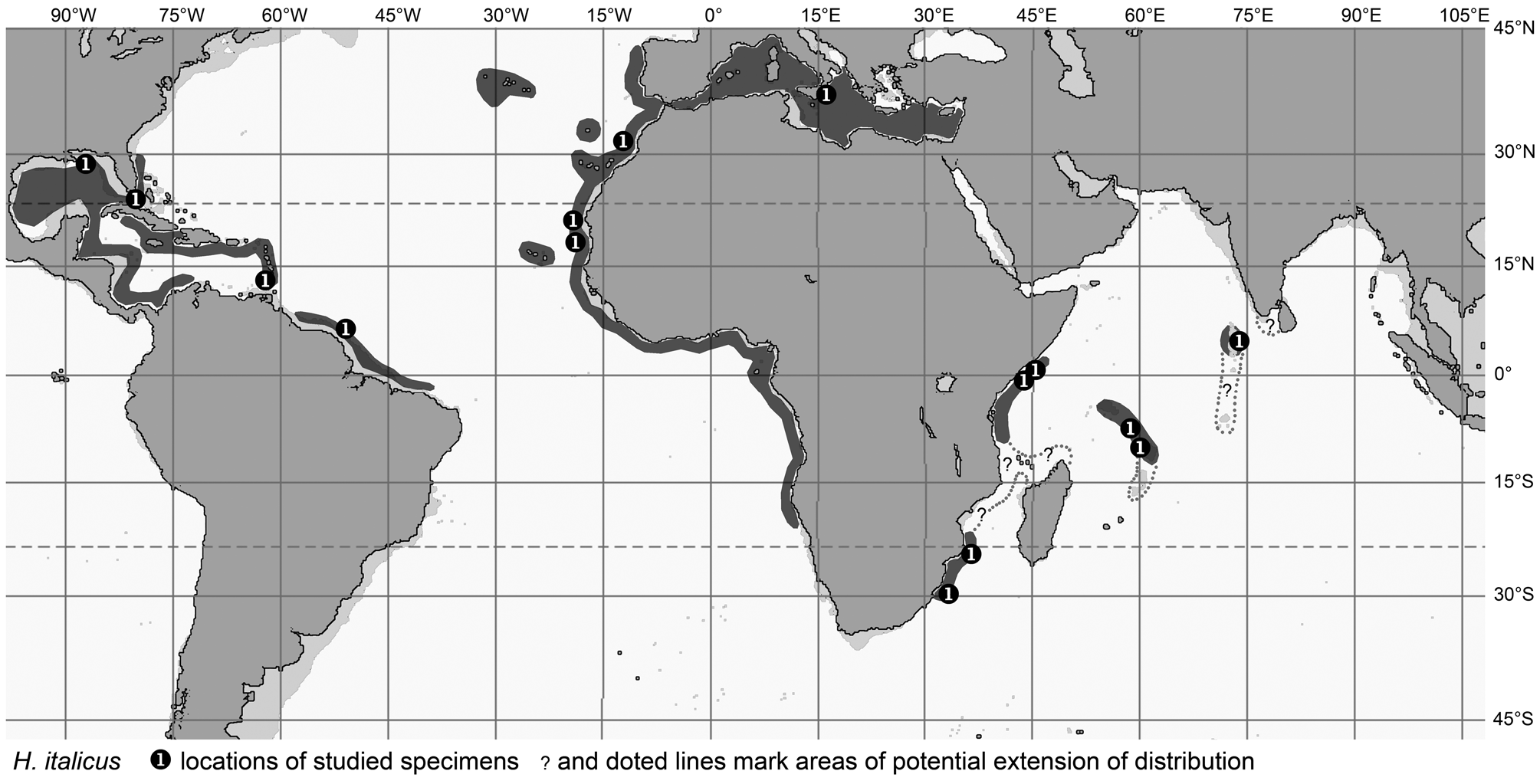
Some other species with relatively wide geographic distribution are H. italicus in the Atlantic and western Indian Ocean, the three closely related species H. aeger , H. striatissimus and H. torvus (considered as of subspecies ranking by Gilbert & Hubbs, 1920), the closely related pair H. lethonemus and H. nascens , and finally H. longibarbis , all in the same general area of the western Pacific, from southern Japan to Australia. Obviously, these species do show some regionalization. In the case of H. italicus there are some very subtle differences in fishes from the western Atlantic, eastern Atlantic and western Indian Ocean, but they are so small and inconsistent that they do not warrant differentiation into species or subspecies at this stage. The same is true for H. longibarbis , but the other two complexes mentioned above do show reasonable morphologically stable differentiation that in both instances led to the recognition of separate taxa in the area of southern Japan to Taiwan, as compared to the region from the Philippines to Australia. In the H. striatissimus complex even a separate species is recognized along the Philippines, viz. H. torvus . The differentiation of the respective species in these two species complexes depends on mostly subtle characters and still carries some tentative elements requiring more specific detailed analyses. Regardless of the outcome of such in-depth statistic analysis and whether differentiation is viewed as of species ranking or subspecies ranking, which appear to be the main alternatives, I think it is fair to assume that the systematic observations already made call for a rather young speciation event. A possible root cause for a geologically young speciation event might be expected in the rapid sea level changes during the Pleistocene and the temporary isolation effects that possibly came with it.
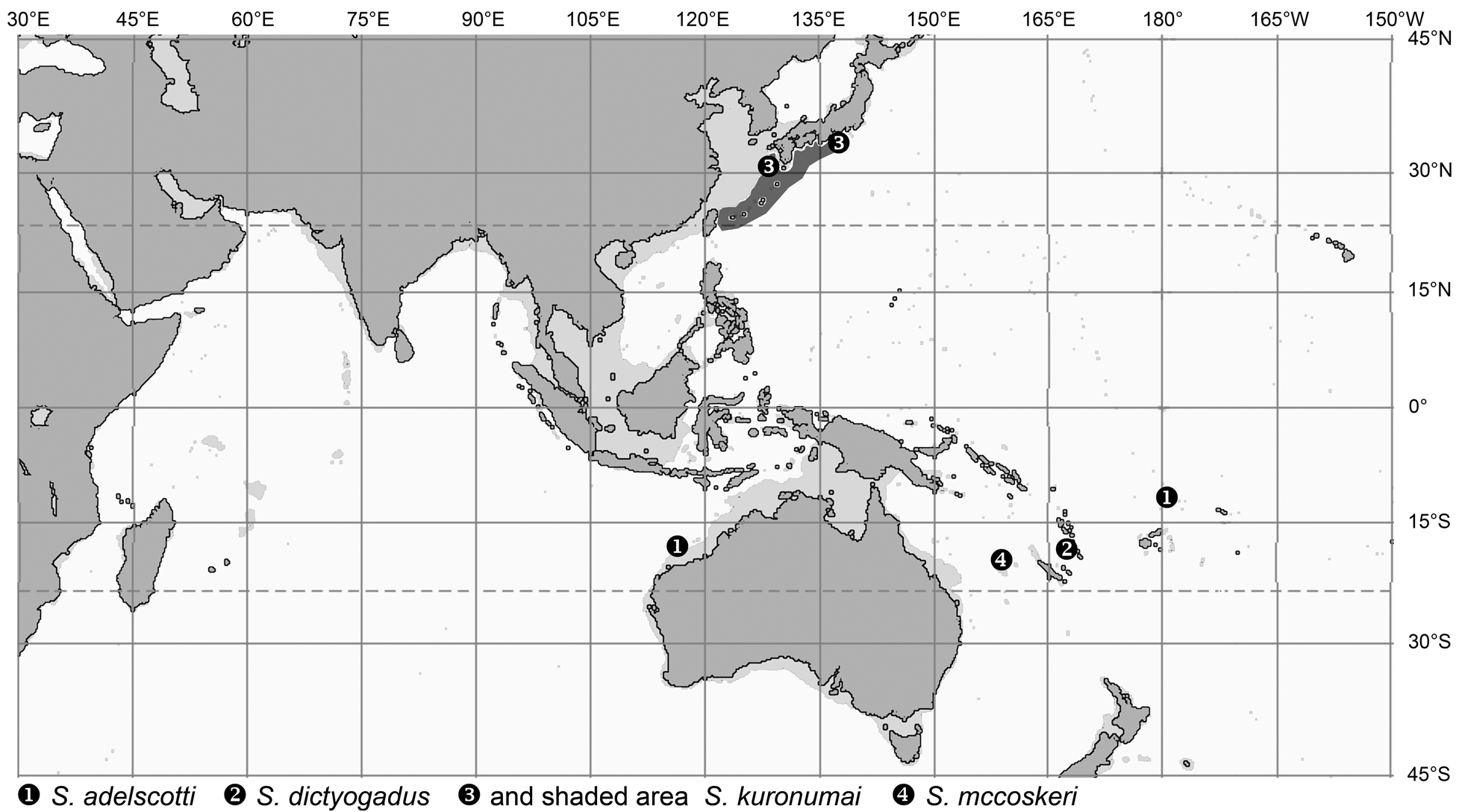
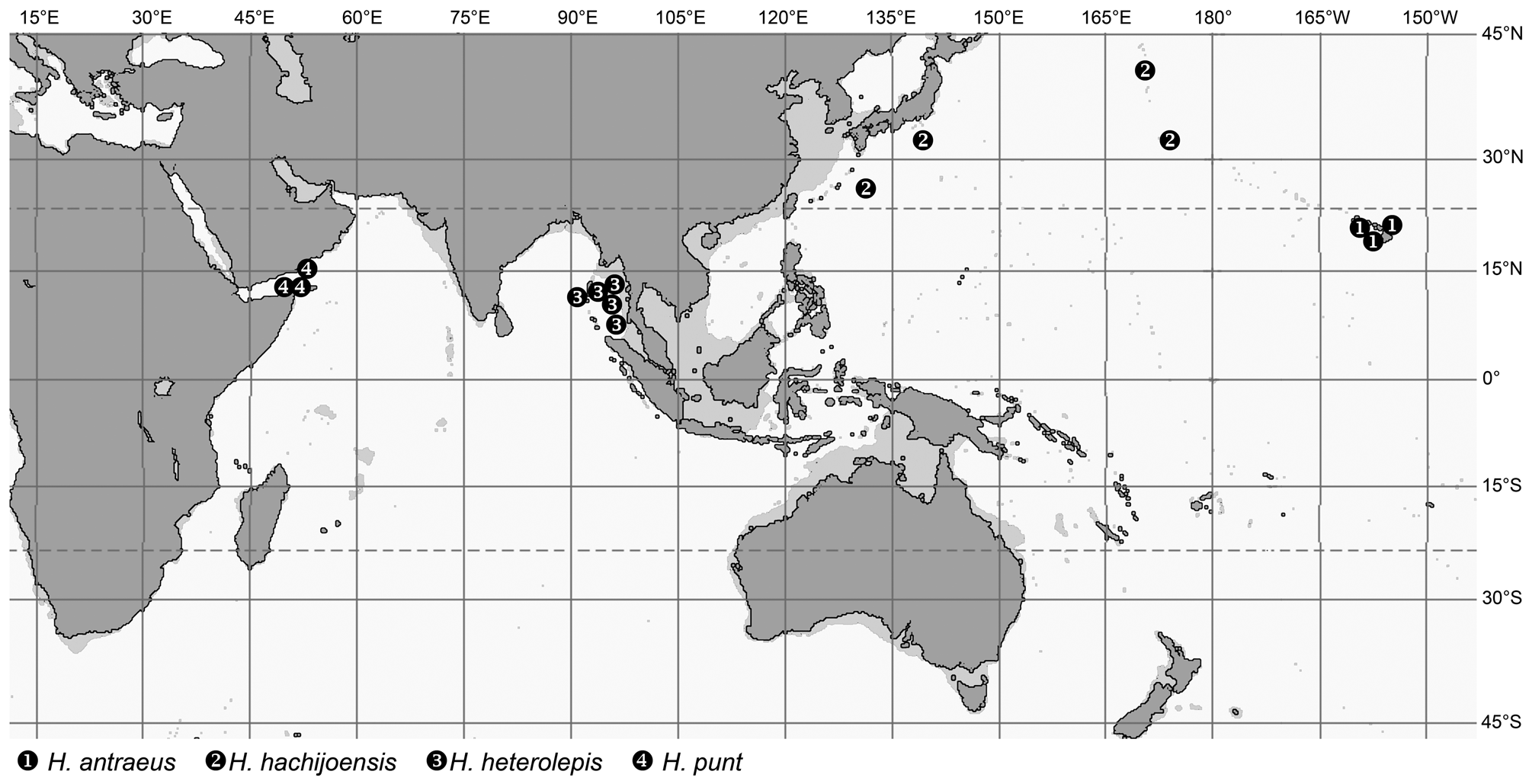
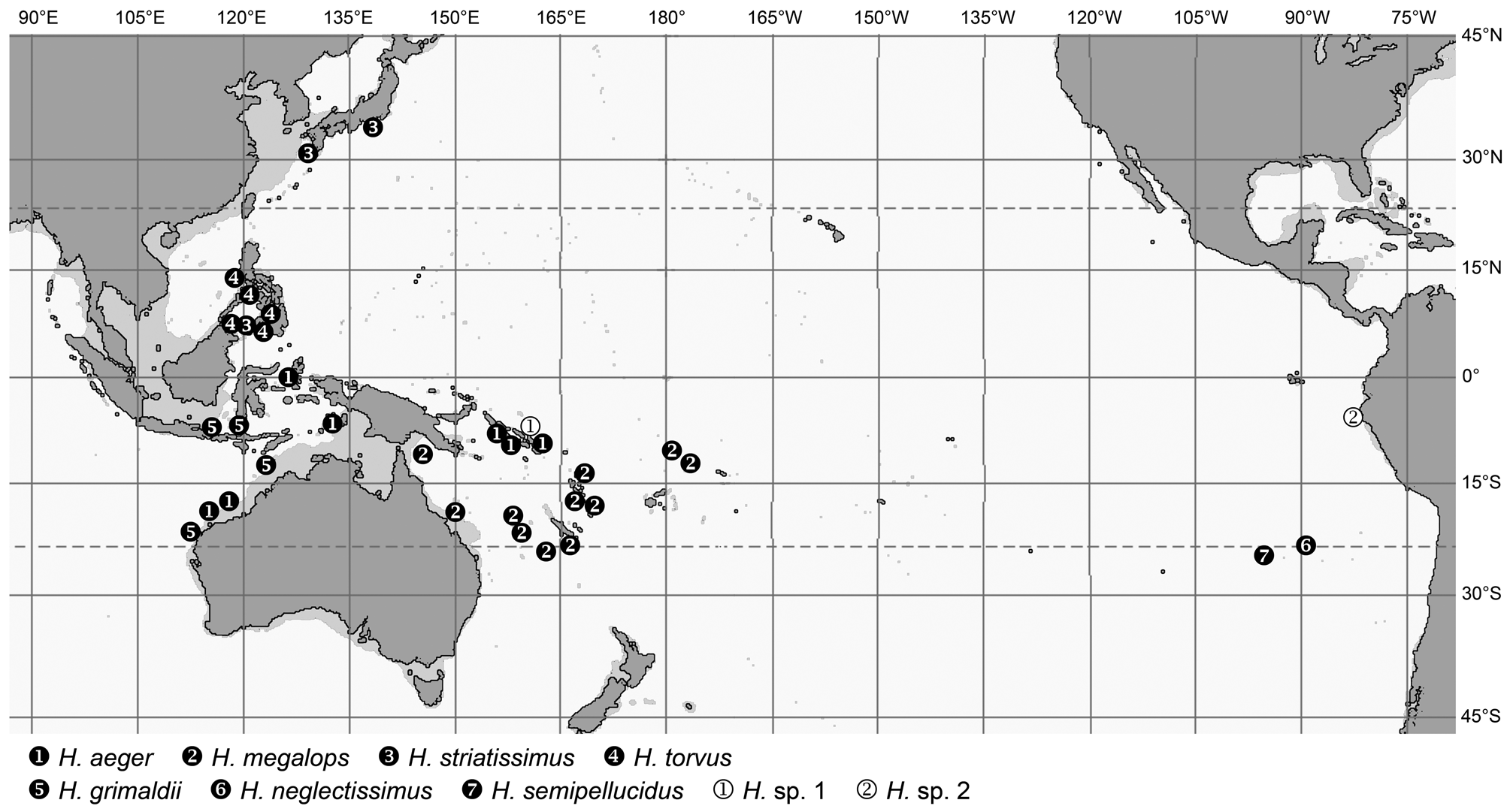
The genera Spicomacrurus , and much more so Hymenocephalus , also include a number of species with seemingly very restricted distributions, i.e. potentially endemic species. These occur in a variety of regions: H. heterolepis in the northeastern Indian Ocean and H. punt chiefly in the Gulf of Aden, but the majority of endemic species are found in the western and central Pacific. The areas richest with endemics are Japan ( Spicomacrurus kuronumai , Hymenocephalus lethonemus , H. papyraceus , H. striatissimus ) and the SW-Pacific from eastern Australia to New Caledonia, Vanuatu and Fiji ( Spicomacrurus dictyogadus , S. mccoskeri , Hymenocephalus megalops , H. nesaeae ). Of these, S. dictyogadus , H. nesaeae and H. papyraceus may be regarded as plesiomorphic species in their respective species groups. The Philippines, Hawaii and Sala y Gomez Ridge all have three species restricted to the specific areas, and Hawaii and Sala y Gomez a fourth species, H. striatulus , which they share. The high degree of endemism in Hawaii and Sala y Gomez reflects ‘outposts’ of the distribution range of the genus Hymenocephalus and most species are considered derived in respect to their species groups (i.e. Hymenogadus tenuis , Hymenocephalus antraeus , H. neglectissimus , H. semipellucidus ), but two others are considered plesiomorphic counterparts, H. sazonovi for H. aterrimus and H. striatulus within their species group. Somewhat unexpectedly the NW Australian continental slope and the adjacent Timor and Java Seas have only yielded two endemic species so far— Hymenogadus grimaldii and H. iwamotoi —both with rather advanced morphologies when compared to their respective sister taxa, and at the same time rather isolated in their systematic position, which suggest a longer isolation time.
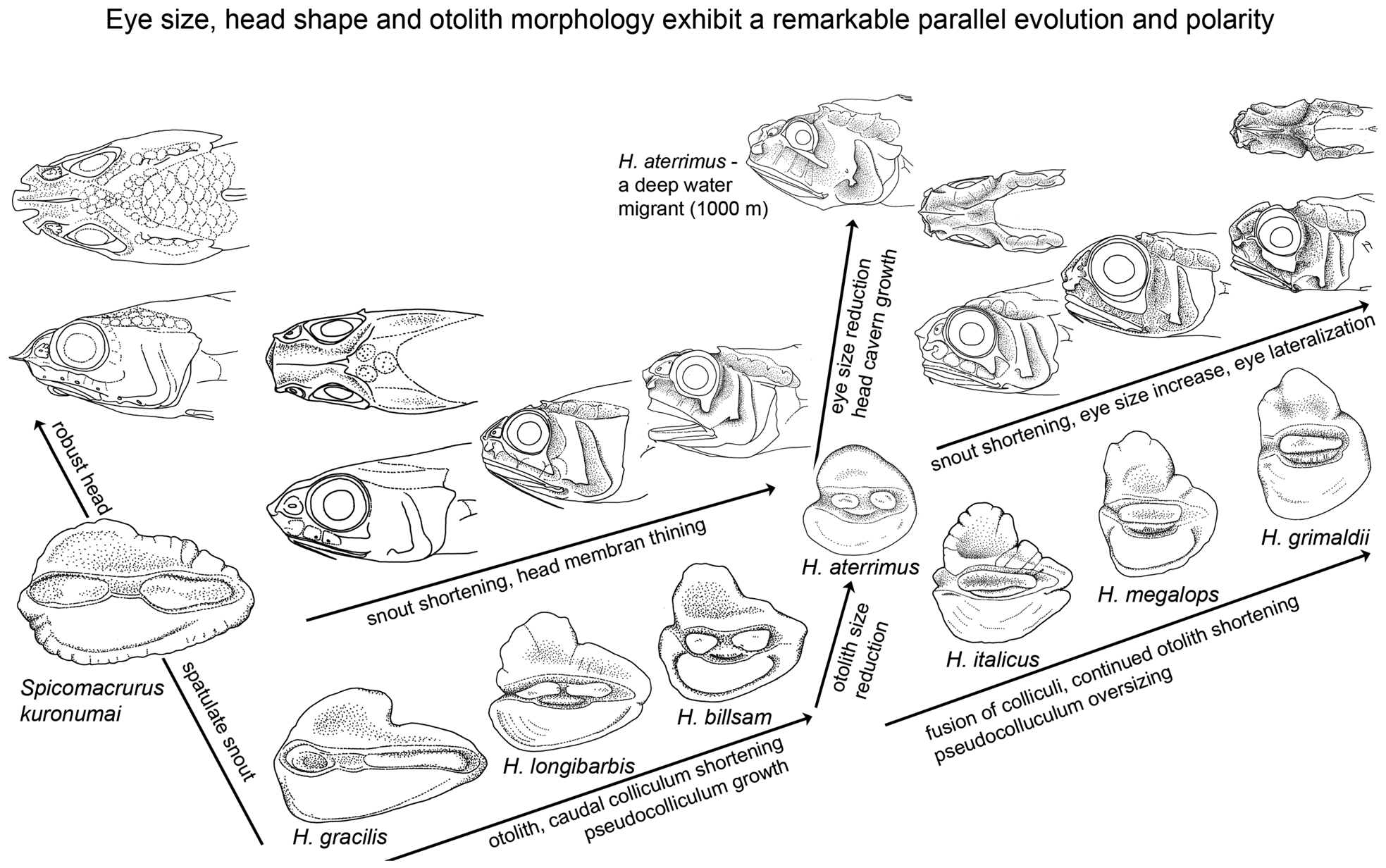
Phylogenetic trends and polarities of characters ( Fig. 39 View FIGURE 39 )
The spectacular development of organs for visual, auditory and motion reception senses within the genus Hymenocephalus likely represents a functional morphological adaptation, which is yet to be fully understood (see also Marshall, 1979, pp. 276–277, 380–383, 413–420, 422–425). Irrespective of what the exact triggers for this evolution might eventually prove to be, it is already now clear that development of the specialization of these three traits follow specific patterns in the genera studied and in large part developed in parallel. In this respect, Spicomacrurus represents an evolutionary trend quite different from Hymenocephalus , with Hymenogadus plesiomorphic to both but probably closer to Hymenocephalus .
The species of Spicomacrurus show a remarkable development of large, robust and spatulate nasal bones not seen in any species of the two other genera and projecting forward above a ventrally flattened head profile. The eyes are oriented at about 60–45° angle to the fish axis resulting in a more dorsally oriented field of vision than the evolution of the visual field in Hymenocephalus . Both are aspects that probably speak for a rather benthic way of live, as already concluded in Okamura (1970). The otoliths do not show any specific specialization. Within Spicomacrurus , S. dictyogadus represents the most plesiomorphic species with the least strongly developed nasal bones and the least flattened ventral head profile. Spicomacrurus mccoskeri is the one with the longest nasal projection and the most dorsally oriented field of vision (indicated by the narrow interorbital ratio of 25% HW).
Three main trends of specialization are observed in the development of the sensory organs in Hymenocephalus .
1. Eye / orbit size and orientation. Orbit sizes range from 20–50% of HL in Hymenocephalus . Orbit sizes in the range of 28–35% of HL are found in species which I consider more basal within the genus, i.e. in the longibarbis , striatulus and antraeus Groups. Nevertheless , these are already fairly large diameters when compared to those of many other macrourids. Orbit sizes in the range of 40–50% of HL are reached in H. antraeus and the striatissimus and grimaldii Groups , whereby the latter two are considered amongst the most advanced in the genus. These observations document a clear polarity of characters related to the visual sense, but there is also a prominent polarity reversal evident in the aterrimus Group with small, supposedly reduced orbit sizes in the range of 20–25% of HL (only exception is H. papyraceus with up to 30% HL). The lateralization of the eyes / orbits is advanced in most species of Hymenocephalus with interorbital values of 50% of HW or more except for the longibarbis Group and H. megalops . The highest values in the range of 70–80% of HW are found in H. grimaldii and H. papyraceus . The large eye size, the ‘parabolic’ shape of the orbit and its advanced lateralization combined with a relatively deep, laterally compressed body shape of the fishes all speak for a way of life detached from the sea bottom in free water, but not pelagic, since Hymenocephalus specimens are usually caught over continental and ocean island slopes and not or very rarely in the open ocean. The polarity reversal in the development of the eye and orbit sizes in the aterrimus Group probably represents an adaptive regression as a result of a somewhat deeper mode of life of these species (generally 700–1000 m) in an area with further reduced light than that for most other species of the genus (generally 300–600 m).
2. Head canals / lateral line system on head. One of the most striking specializations found in the genus Hymenocephalus is the development of very spacious and deep head canals housing the lateral line system on the head and its rarely preserved thin skin cover (‘integument’ or ‘hymen’) spanned over the canals with the help of supporters. This specialization is not seen in Spicomacrurus and only incipiently in the outgroup constituted by Hymenogadus . Not much is to be recognized within Hymenocephalus in terms of polarity of character development related to head canals. The two species of the longibarbis Group are remarkable for their relatively narrow supraorbital cavern width (6–10% vs 9–20% HL in other species of the genus, except 5–8% HL in H. neglectissimus and H. semipellucidus ), and a relatively wide postorbital-preopercular interspace (10–14% vs 4–10% HL in most other species of the genus, except 10–14% in H. aterrimus and H. sazonovi ). Both character states are considered plesiomorphic within the genus and hence support the basal phylogenetic position of the longibarbis Group. The one species group sticking out from the remainder of the genus is the aterrimus Group, which shows the widest head canals of all Hymenocephalus species : infraorbital width 16–23% HL (vs 6–17% HL), supraorbital width mostly 13–20% HL (vs 9–16, except up to 18% HL in H. grimaldii ) and preopercular cavern width mostly 15–22% HL (vs 8–16%, except up to 19% HL in H. grimaldii ). Again, the size increase of the head canal system observed in the aterrimus Group might be an adaptation to their way of life in deeper waters when compared to other species of the genus, with probable orientation towards motion-capturing senses at the expense of visual and hearing capacities (see above and below).
3. Sagittal otolith size and morphology. Four individual trends of specialization are observed in sagittal otoliths: the increase of relative size to more than 20% HL in some species ( Fig. 2A View FIGURE 2 ), the foreshortening of the shape with an enormously large and high predorsal lobe, the development of an extremely long pseudocolliculum in the ventral part of the collum extending well beyond the ostial and caudal colliculi, and the fusion of the ostial and caudal colliculi. In all these aspects otoliths of the genus Hymenogadus , representing the outgroup for Hymenocephalus , show a plesiomorphic character state. The otolith morphology found in the longibarbis Group shows a transitional status, advanced in comparison to Hymenogadus , but basal in respect to all other species groups of Hymenocephalus except for the aterrimus Group. In fishes of the aterrimus Group, otolith sizes are regressed between 10–15% HL, being smallest in H. aterrimus at about 10–11% HL. The pseudocolliculum is still short (TCL:PCL = 1.8–2.8), the outline is compressed (OL:OH = 0.8–1.2), and the predorsal lobe well developed except for H. aterrimus where it is shallow and rounded, probably as an indication of reversed polarity, like the overall reduction of otolith size. The reduction of otolith size appears to be related to life in deeper waters, as demonstrated by Wilson (1985) for several macrourid fishes, and sometimes observed within the same species caught at different water depths. The compression of the otolith shape shows a character polarity from an OL:OH ratio of 1.1–1.3 in the longibarbis Group to 0.75–0.8 found in the striatissimus and grimaldii Groups. The relative length of the pseudocolliculum shows a character polarity from a ration TCL:PCL of 1.7–2.8 in the longibarbis , iwamotoi , aterrimus and billsam Groups to a ratio of less than 1.35 found in the grimaldii Group and certain species of the striatissimus Group. Otoliths with a similar outline are found in other macrourid genera as well, for instance in Squalogadus , Macrouroides and Trachyrincus ( Nolf & Steurbaut, 1989) , but, as stated before, the very large size of the pseudocolliculum is a unique character so far observed in teleost otoliths. The fusion of the ostial and caudal colliculi finally is observed in the italicus , striatissimus and grimaldii Groups and incipiently though only occasionally in H. antraeus . The fusion of the colliculi into a single undivided feature is commonly observed in benthic and benthopelagic fishes and certainly reflects a case of functional morphological adaptation, which is not yet understood. A number of studies exist that evaluate the pattern of the hair cell orientation of nerve ciliae attached to the sulcus of the sagittal otolith; these show great variation, but the effect on the otolith or sulcus morphology is not yet understood. The only gadiforms seemingly studied in this respect are bregmacerotids ( Popper, 1980), merlucciids ( Lombarte & Popper, 2004) and morids ( Deng et al., 2011), but no macrourid. Schwarzhans (1981) has shown the occurrence of multiple, unrelated developments of fused colliculi in the Ophidiiformes , sometimes within a genus, similar to that in Hymenocephalus . Similar observations no doubt could be made with otoliths of the Macrouridae , but in Hymenocephalus there is no indication that fusion of the colliculi has occurred more than once.
In summary, it appears that the specializations of characters associated with the visual, auditory and motioncapturing senses (eye / orbit size, sagittal otolith morphology and head canal system) have developed in parallel and along similar polarities within the genus Hymenocephalus , with the longibarbis Group the most basal and the striatissimus and grimaldii Groups the most advanced ( Fig. 39 View FIGURE 39 ). Polarity reversal is observed in two of the three main trends (visual and auditory) in the aterrimus Group, which is probably a result of adaptation to a life in deeper waters.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |