
Hymenocephalus sazonovi, Schwarzhans, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3888.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:1B437AE1-CF28-4C1B-95B6-C31A295905A0 |
|
persistent identifier |
https://treatment.plazi.org/id/463A8F36-FF95-FFDF-1297-9B50E3CBFDC5 |
|
treatment provided by |
Felipe |
|
scientific name |
Hymenocephalus sazonovi |
| status |
sp. nov. |
Hymenocephalus sazonovi View in CoL n.sp.
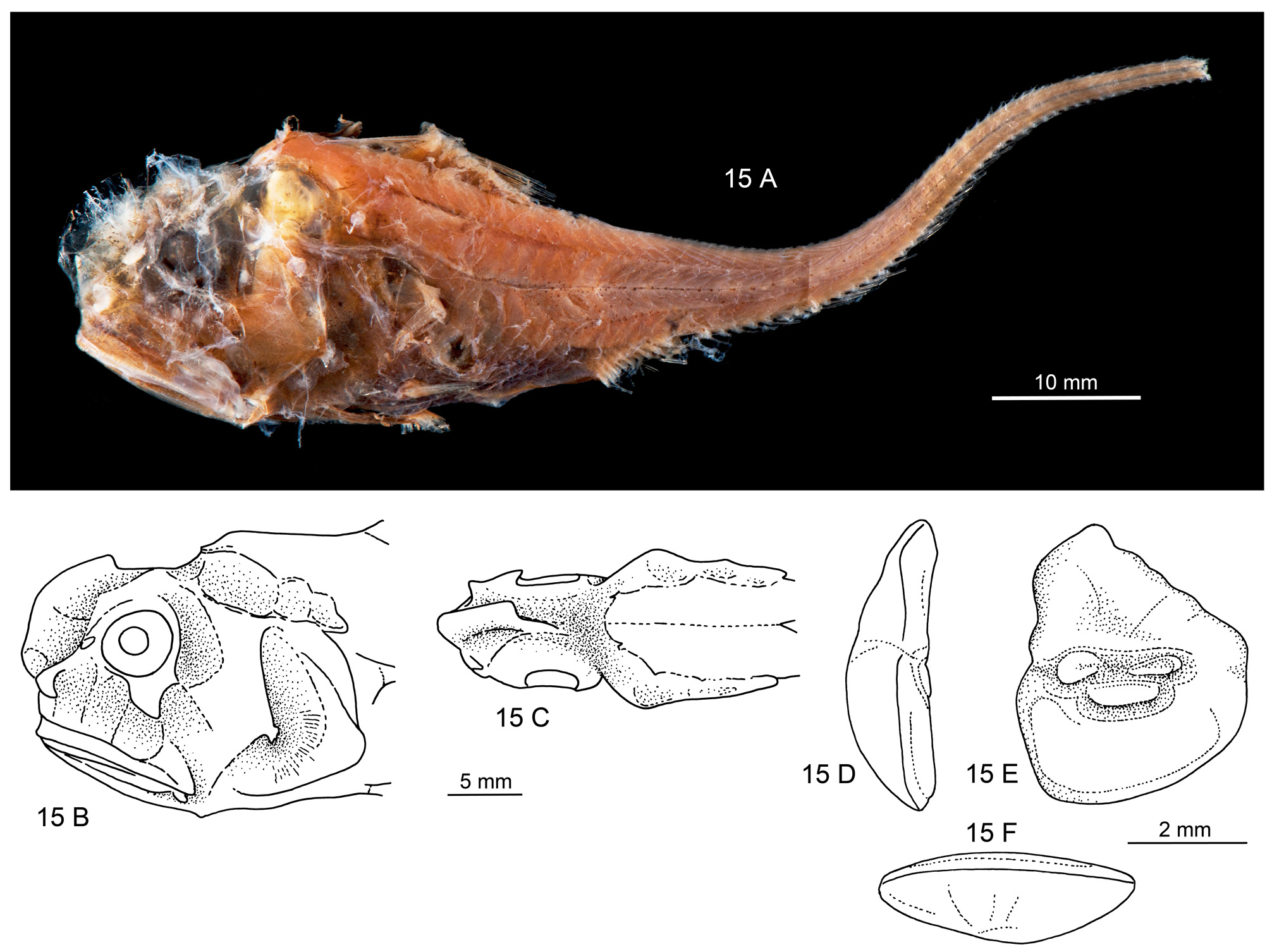
Figs. 15A–F View FIGURE 15 , 16 View FIGURE 16
Hymenocephalus sp. cf. aterrimus: Sazonov & Iwamoto, 1992: 54 View in CoL .
Material examined (3 specimens, 84+ –131+ mm TL). Holotype ZMMGU P-18128, 84+ mm TL, Nazca Ridge, 340–780 m, Prof. Mesiatzev cruise 13 ; Paratypes: ZMMGU P-17729, 121–131+ mm TL, Sala y Gomez Ridge, 1070–1100 m, Prof. Mesiatzev cruise 13, trawl 10 .
Diagnosis. A stout species, much less slender than typical for the genus, with very thin head bones. Pelvic fin rays 12–13; pectoral 15–16; gill rakers 18–20. Barbel absent. Orbit 21% HL. Snout blunt, with high frontal ridge, barely protruding, short, 23% HL. Ventral striae extending about ⅔ distance from pelvic fin bases to periproct region. Infraorbital canal wider than other canal systems; preopercular supporter very small. Otolith compressed (OL:OH = 0.8); colliculi separated, small; pseudocolliculum small (TCL:PCL = 1.8).
Comparison. Hymenocephalus sazonovi no doubt is closely related to the widespread H. aterrimus Gilbert, 1905 , with which it was tentatively associated when first described by Sazonov & Iwamoto (1992). It differs from H. aterrimus in the deeper head and the high frontal ridge, as already noted by Sazonov & Iwamoto, and the lower number of gill rakers on the inner side of the first gill arch and the outer side of the second gill arch, respectively (18–20 vs 21–27) and the high-bodied otolith with a strongly developed predorsal lobe, which is strongly reduced and rounded in H. aterrimus . This additional otolith character particularly supports the recognition of a separate species.
Description. Counts (from 2 specimens): 1D. II,9 (II,9–10); P. 16 (15–16); V. 13 (12–13); first gill raker inner side 20 (18–20); second gill raker outer side 20 (19–20).
Measurements (from holotype, supplemented in part from one paratype and from Sazonov & Iwamoto, 1992): head length 26.5–35.0 mm, about 25–30% TL; head height 85% HL; head width 50% HL; snout 23% HL; orbit 21% HL; postorbital 56% HL; interorbital 26% HL / 54% HW; upper jaw length 51% HL; pre-anal about 145% HL; pre-dorsal to 1D about 95% HL; pre-dorsal to 2D about 165% HL; distance base 1D to 2D about 70% HL; 1st dorsal fin length about 40% HL; ventral fin length at least 30% HL; supraorbital canal width 15.5% HL; infraorbital width 20% HL; minimal infraorbital canal width 13% HL.
The following description is based on the holotype ( Fig. 15A View FIGURE 15 ). Body stout with massive head, highest and widest just behind orbit, tapering rather abruptly between first dorsal and second dorsal into the usual whip-like tail. Origin of first dorsal, pectoral and ventral fins about on same vertical. Pectoral and ventral incomplete, probably rather short. Second dorsal rudimentary; anal well developed.
Teeth all small, on narrow bands of both jaws.
Luminescent tissue (striae) silvery and well visible as a broad band along both sides of isthmus below gill covers, reaching up to bases of pectoral fins, then turning darker and less clearly visible behind ventral fin bases, where they join along medial line, and expanding to about ⅔ from ventral fin bases to periproct region. Several distinct, broad dark striae on gular region. Anterior lens of ventral luminescent organ small; posterior lens before periproct about two times larger, tear-drop shaped, anteriorly with triangular stretch of dark tissue.
Axial skeleton (based on radiographs). Number of precaudal vertebrae 10; vertebrae 1 to 3 much shorter than subsequent vertebrae. Neural spines of vertebrae 1 and 2 about 30% longer than vertebra 3; neural spines 3 to 7 or 8 depressed and with blunt tips; neural spines 3 to 6 short and of equal length, spines 7 to 8 increasing in length. Bases of neural spines 4 to 8 enlarged. Parapophyses on vertebrae 6 or 7 to10. Pleural ribs on vertebrae 4 or 5 to 8. First fin ray of 1D supported by two pterygiophores, both inserted behind neural spine 2. Last pterygiophore of 1D inserted behind neural spine 7 or 8. First pterygiophore of 2D above vertebra 18 to 20. First pterygiophore of anal fine not prolonged, inserted in front of first haemal spine on first caudal vertebrae (11).
Head morphology ( Fig. 15B–C View FIGURE 15 ): Head stout, with blunt anterior profile, snout barely protruding beyond mouth, with high frontal ridge. Bones very thin and fragile; eyes very small; barbel absent. Head canals well developed, covering integument not preserved in any of the studied specimens. Infraorbital canal wider than all other canal systems (20–22% HL), supraorbital canal with 4 segments, width 13–14% HL, supratemporal canal not identified, preopercular canal 15% HL, postorbital-preopercular interspace 11–12% HL. Infranasal supporter small; infraorbital supporter expanded only below rear part of orbit, about 50% OD; preopercular supporter very small (about 3% HL), forked; mandibular hook small.
Otolith morphology ( Fig. 15D–F View FIGURE 15 ): Otolith high bodied; OL:OH = 0.8; OL:OT about 2.5. Predorsal lobe massive, regularly dropping down distally to broadly rounded posterior tip. Anterior rim irregular, nearly vertical; ventral rim deep, smooth and regularly curved, deepest anterior of middle. Inner face slightly convex along horizontal axis, with narrow, slightly supramedian sulcus. Ostial and caudal colliculi small, narrow, particularly the caudal colliculum; pseudocolliculum moderately enlarged. CCL:OCL = 1.2; TCL:PCL = 1.8. Dorsal depression indistinct; ventral furrow distinct, moderately close to ventral rim.
Coloration (in alcohol) ( Fig. 15A View FIGURE 15 ): Dark brown to black, darkest over trunk and ventral region in front of periproct; tail dark brown. Some black melanophores on head, particularly opercle and along a midlateral band beginning above and behind pectoral fin base.
Distribution ( Fig. 16 View FIGURE 16 ). Probably endemic to the Nazca and Sala y Gomez ridges.
Etymology. The new species is named in memory of the late Yuri I. Sazonov (Moscow, Russia) in recognition of his many contributions to the knowledge of the family Macrouridae . He was also the one first to record the specimens, together with T. Iwamoto, in 1992.

| ZMMGU |
Zoological Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hymenocephalus sazonovi
| Schwarzhans, Werner 2014 |
Hymenocephalus sp.
| Sazonov, Y. I. & Iwamoto, T. 1992: 54 |