
Careproctus faunus, James Wilder Orr & Katherine Pearson Maslenikov, 2007
|
publication ID |
https://doi.org/ 10.1643/0045-8511(2007)2007[699:Tnvsot]2.0.Co;2 |
|
DOI |
https://doi.org/10.5281/zenodo.5611919 |
|
persistent identifier |
https://treatment.plazi.org/id/46309522-2C0F-FFC2-B9B4-FE620550FD60 |
|
treatment provided by |
Plazi |
|
scientific name |
Careproctus faunus |
| status |
sp. nov. |
Careproctus faunus View in CoL , new species
Mischievous Snailfish
Figures 1B, 2B, 3, 4; Tables 1 View TABLE 1 , 2 View TABLE 2
Holotype.— UW 111867 , 101.9 mm, male, Amukta Pass , 52.21°N, 172.21°W, 348 m depth, F/V SEA STORM, cruise 200201, haul 157, 23 July 2002, J. W. Orr. GoogleMaps
Paratypes.— 41 specimens, 24.0– 101.9 mm. UW 113667, 2, 58.6–68.7 mm, 52.28° N, 172.94° W, 291 m depth, F / V VESTERAALEN, cruise 200001, haul 65, benthic bag, 3 June 2000; UW 113671, 91.3 mm, female, collected with holotype; UW 111835, 3, 73.8–85.8 mm, 52.85° N, 170.57° W, 329 m depth, F / V DOMINATOR, cruise 200001, haul 37, benthic bag, 26 May 2000, J. W. Orr; UW 111837, 63.7 mm, 52.38° N, 170.66° W, 278 m depth, F / V VESTERAALEN, cruise 200001, haul 51, benthic bag, 31 May 2000; UW 111839, 69.0 mm, 52.65° N, 172.26° W, 376 m depth, F/ V DOMINATOR, cruise 200001, haul 84, benthic bag, 11 June 2000, K. E. Pearson; USNM 387975, 2, 62.4–77.0 mm, 52.99°N, 169.89°W, 120 m depth, F/V VESTERAALEN, cruise 200001, haul 36, benthic bag, 27 May 2000, W. C. Flerx; UW 111857, 90.2 mm, 52.33°N, 172.75°W, 444 m depth, F/V VESTERAALEN, cruise 200001, haul 67, 4 June 2000; UW 111868, 5, 80.5–96.5 mm, 52.37°N, 171.24°W, 323 m depth, F/V DOMINATOR, cruise 200001, haul 51, benthic bag, 31 May 2000, J. W. Orr; CAS 224213, 2, 73.0– 79.9 mm, 51.62°N, 176.25°W, 321 m depth, F/V VESTERAALEN, cruise 200201, haul 191, 17 July 2002; SIO 06-108, 2, 74.0–90.0 mm, 52.94°N, 169.44°W, 422 m depth, F/V MORNING STAR, cruise 200201, haul 43, benthic bag, 2 June 2002, J. W. Orr; UW 113639, 56.0 mm, 53.15°N, 168.99°W, 259 m depth, F/V MORNING STAR, cruise 200201, haul 24, 30 May 2002, J. W. Orr; UW 113640, 2, 85.0– 88.8 mm, 52.34°N, 170.65°W, 226 m depth, F/V SEA STORM, cruise 200201, haul 188, 1 Aug. 2002, R. C. Harrison; UW 113641, 85.0 mm, 52.37°N, 171.36°W, 324 m depth, F/V SEA STORM, cruise 200201, haul 225, 10 Aug. 2002, R. C. Harrison; UW 113644, 76.7 mm, 52.41°N, 170.62°W, 313 m depth, F/V SEA STORM, cruise 200201, haul 198, 3 Aug. 2002; UW 113645, 78.2 mm, 53.12°N, 169.94°W, 219 m depth, F/V MORNING STAR, cruise 200201, haul 29, benthic bag, 30 May 2002, J. W. Orr; UW 113646, 2, 90.0–96.0 mm, 52.21°N, 172.21°W, 341 m depth, F/V SEA STORM, cruise 200401, haul 73, 23 June 2004, J. W. Orr; UW 113647, 72.1 mm, 51.76°N, 175.46°W, 272 m depth, F/V VESTERAALEN, cruise 200201, haul 60, benthic bag, 11 June 2002, R. N. Clark; UW 113651, 66.0 mm, 54.18°N, 166.34°W, 266 m depth, F/V VESTERAALEN, cruise 200201, haul 31, benthic bag, 28 May 2002; UW 113653, 82.6 mm, 52.95°N, 169.31°W, 397 m depth, F/V GLADIATOR, cruise 200401, haul 30, benthic bag, 14 June 2004, K. E. Pearson; UW 113658, 3, 48.3– 75.5 mm, 52.83°N, 170.56°W, 332 m depth, F/V MORNING STAR, cruise 200201, haul 50, benthic bag, 3 June 2002, J. W. Orr; UW 113662, 60.0 mm, 52.78°N, 170.69°W, 324 m depth, F/V DOMINATOR, cruise 200001, haul 38, benthic bag, 26 May 2000, J. W. Orr; UW 113663, 2, 24.0– 72.2 mm, 52.55°N, 170.11°W, 230 m depth, F/V SEA STORM, cruise 200401, haul 32, 14 June 2004, J. W. Orr; LACM 56483–1, 2, 82.2–82.9 mm, 52.39°N, 170.67°W, 281 m depth, F/V SEA STORM, cruise 200201, haul 187, benthic bag, 1 Aug. 2002, R. C. Harrison.
Diagnosis.— Careproctus faunus is diagnosed from all other described species of Careproctus by its body coloration of red and gray mottling. It is further distinguished from Careproctus comus , new species, by the following character states: dorsalfin rays 47–51 (vs. 50–56 in C. comus ); anal-fin rays 41–45 (vs. 44–50); caudal vertebrae 42–45 (vs. 45–51); interorbital space wider, 11.9–16.7 % HL (vs. 7.0–13.6 % HL); maxilla longer, 37.4– 49.0 % HL (vs. 32.0–46.0 % HL); snout longer, 20.1–34.9 % HL (vs. 16.9–28.2 % HL); connection of anal-fin membrane to caudal fin longer, 34.8– 56.3 % CL (vs. 26.7–45.6 % CL); caudal-fin base deeper, 11.7–19.7 % CL (vs. 10.0–17.2 % CL).
In addition, the following combination of character states distinguish C. faunus from all other species of Careproctus : snout protruding beyond lower jaw; pelvic disk moderate in size, 6.9–9.3 %; pectoral fin with shallow notch and short lower lobe; distal pectoral radials absent from base of 8–12 ventral rays; teeth trilobed.
Description.— Body slender, tapering strongly posteriorly, moderately compressed; depth at dorsalfin origin 60–100 (75.7) % HL. Head moderately large, dorsal profile gradually sloping from nape to snout. Snout rounded, strongly projecting beyond upper jaw. Mouth small, maxilla 37.4– 49.0 (40.8) % HL, extending to mid-orbit, oral cleft extending to anterior rim of orbit. Lower jaw inferior, premaxillary tooth plates matching mandibular tooth plates. Premaxillary teeth trilobed in 9–10 oblique rows, increasing from 3–5 teeth in anteromedial rows to 12 teeth in posterolateral row. Mandibular teeth trilobed in 9–10 oblique rows, increasing from 3–5 teeth in anteromedial rows to 14 teeth in posterolateral rows. Diastema absent at symphysis of upper and lower jaws. Orbit large, diameter 31.7–46.0 (33.0) % HL, dorsal margin below dorsal contour of head, suborbital depth to oral cleft 21.8–57.1 (43.3) % orbit length; pupil elliptical to round. Interorbital space narrow, 11.9–16.7 (14.2) % HL, slightly convex. Snout shorter than orbit, 16.1– 34.9 (27.7) % HL. Nostril single, in well-developed tube at level with middle or upper part of orbit; nostril tube length 2.0–4.0 (3.4) % HL.
Pores of cephalic lateralis of moderate size: nasal pores two, maxillary pores six, preoperculomandibular pores seven, suprabranchial pores two (pore pattern 2-6-7-2); chin pores paired in separate pits. Interorbital pore absent. Free neuromasts ( Andriashev and Stein, 1998) about 5–6, small and difficult to discern, originating from above gill slit and extending to a level at mid-body to about level with anus.
Gill opening small, 16.7–27.0 (25.8) % HL, upper margin at level of mid-orbit or dorsal part of orbit, extending to just above pectoral fin. Opercular flap rounded or angular. Branchiostegal rays six.
Dorsal-fin rays 47–51 (48; Table 1 View TABLE 1 ), anterior 4– 5 rays uniserial and unsegmented, more posteri- or rays biserial and segmented; all rays simple. Anteriormost dorsal-fin pterygiophore inserted between neural spines two and three, bearing a single ray.
Anal-fin rays 41–45 (43; Table 1 View TABLE 1 ), anterior ray uniserial and unsegmented, more posterior rays biserial and segmented; all rays simple. One or two anal-fin pterygiophores each bearing a single ray anterior to first haemal spine. Anal-fin origin below vertebrae 11–12 (caudal vertebrae 1–2).
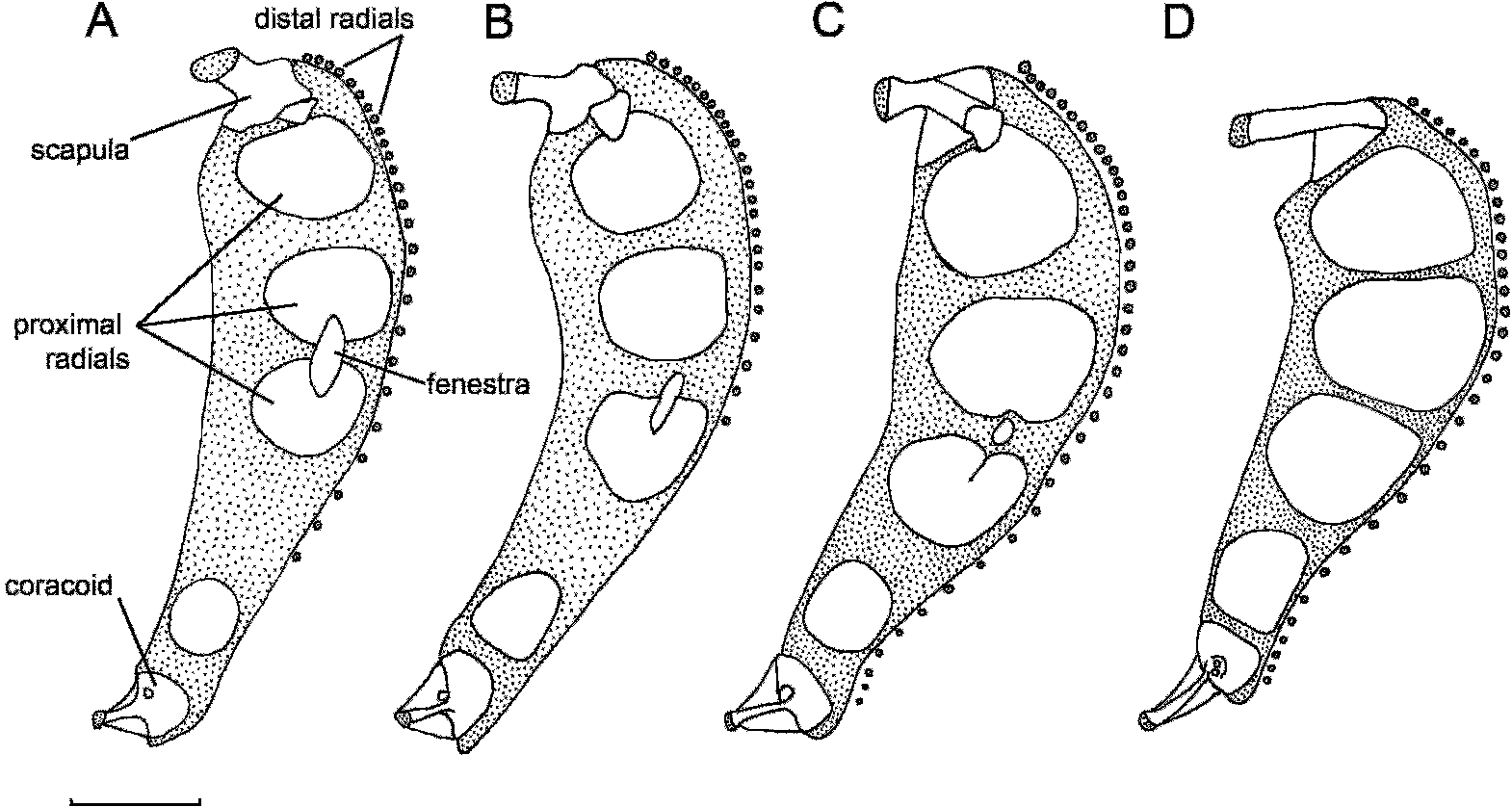
Pectoral fin shallowly notched, with 32–38 (35) rays ( Table 1 View TABLE 1 ). Upper lobe of 25–31 (25) rays extending to anal-fin origin or beyond to anal-fin ray five, dorsalmost rays lengthening to rays 3–4, more ventral rays gradually shortening to shortest ray of notch. Lower lobe short, with 7–10 (10) rays, extending to or slightly beyond anus; dorsal rays gradually lengthening to thick and fleshy rays 2–4, more ventral rays more slender and gradually shortening to ventralmost ray near pectoral symphysis. Tips of rays 5–20 % free of membrane, lower rays more strongly exserted. Rays in notch slightly more widely spaced than rays of lobes. Uppermost pectoral-fin ray level with ventral rim of orbit. Lowermost pectoral-fin ray below anterior part of orbit. Proximal pectoral radials four (3 + 1), robust; proximal radials 2–3 notched ( Fig. 2 View Fig. 2 B). Interradial fenestra one, shape variable: slender to elliptical, formed as a notch in proximal radial two only or extending partially or completely from radial two to three. Scapula with strong helve; coracoid triangular with broad lamina. Distal radials present at base of pectoral-fin rays one to 22– 30; more ventral distal radials reduced, absent from base of ventralmost rays, which articulate directly with pectoral cartilage.
Pelvic disk moderate in size, 24.6–33.0 (30.0) % HL, round, slightly longer than wide, anterior lobe moderately developed or obsolete, flat with margins often slightly upturned. Anus closer to pelvic disk than to anal-fin origin.
Principal caudal-fin rays 11–12 (11; Table 1 View TABLE 1 ), dorsal procurrent rays 2–3 (2), ventral procurrent rays 1–2 (1). Membrane of posterior dorsalfin rays overlapping caudal fin 15.9–43.7 (28.1) % CL; posterior anal-fin rays, 20.3–56.3 (47.5) % CL.
Skin thin, fragile, prickles absent. Pyloric caeca 16–18, thick, length about 25.3–41.5 (26.2) % HL.
Vertebrae 52–56 (53), 9–11 (10) precaudal, 42–45 (43) caudal ( Table 1 View TABLE 1 ). Pleural ribs present on vertebrae 8–10, anteriormost short and slender, those more posterior long and slender. Hypural plate composed of dorsal and ventral plates divided by deep split about 75 % length of plate. Single epural present.
Largest specimen examined a 101.9 mm male (UW 111867, holotype). Smallest female with yolked eggs 91.3 mm (UW 113671, collected with holotype); smallest male with enlarged, swollen testes 58.6 mm (UW 113667).
Coloration in life.— Body pale to dusky, mottled red and white anteriorly, mottled red and gray posteriorly, posterior fins often edged with black ( Fig. 1 View Fig. 1 B). Head with red band extending from tip of snout to anterior rim of orbit, red mottling on cheek and operculum, with bright white blotches along lower jaw and edge of operculum from near tip of jaw to gill slit; lips unpigmented. Upper pectoral-fin rays bright red; middle rays unpigmented proximally, lighter red distally, with bright patch of white anteriorly at middle part of rays; rays of lower lobe white. Caudal-fin rays dusky to light red, often edged with black. Peritoneum and orobranchial cavity pale; stomach, intestines, pyloric caeca, and urogenital papilla pale.
Distribution.— Careproctus faunus has been collect- ed only in the central and eastern Aleutian Islands, from Adak Island (176.3°W) to just east of Unalaska Island (166.3°W) at depths of 120– 422 m ( Fig. 3 View Fig. 3 ).
Etymology.— Named for the Roman god Faunus , the mischievous half-brother of Comus , because of the close similarity of this species to C. comus .
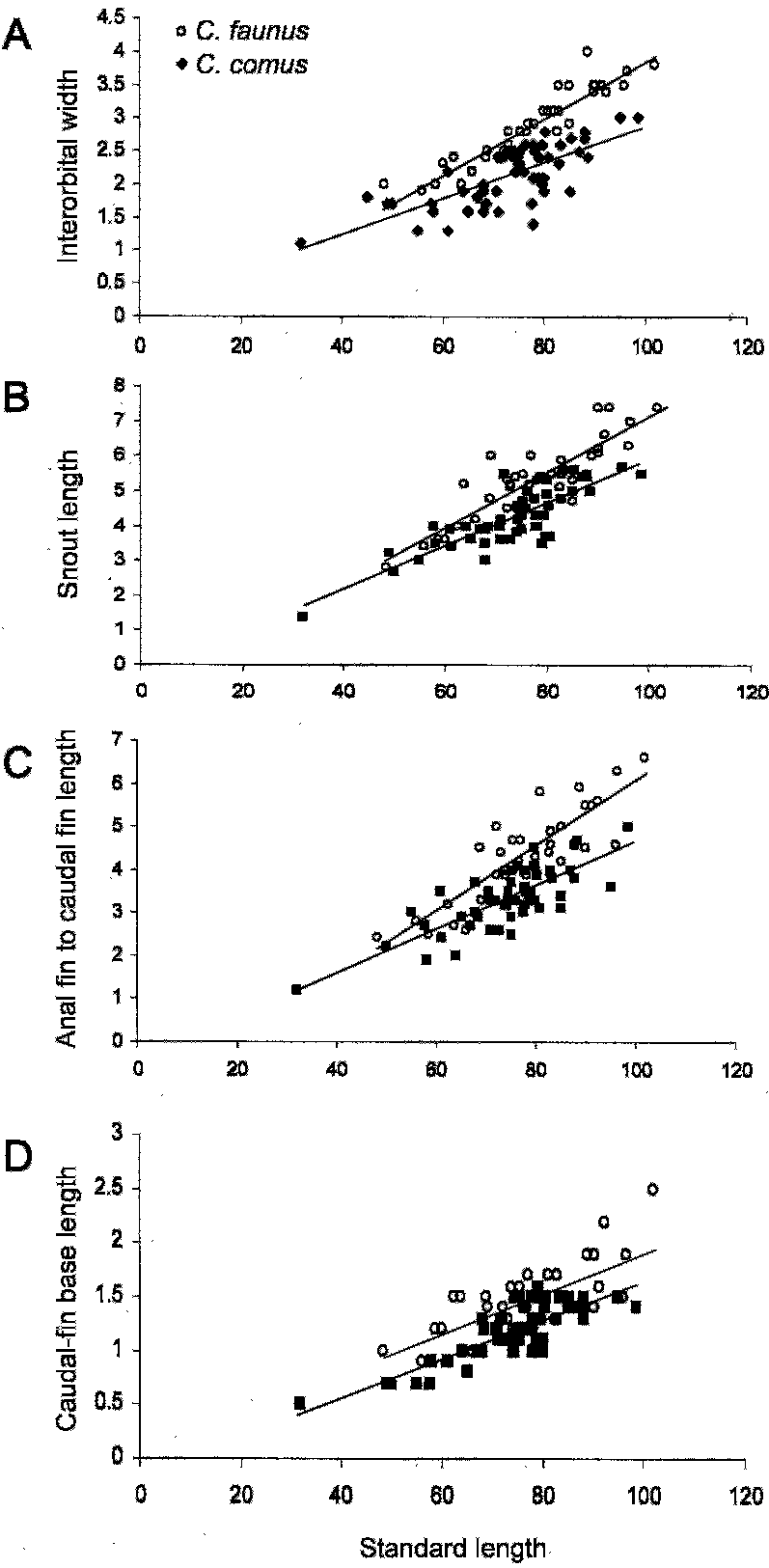
Comparisons.— The two new species, Careproctus comus and C. faunus , are most similar to one another and are apparently closely related. Several statistically significant differences were found between the new species ( Table 1 View TABLE 1 ). Dorsal- and anal-fin rays, and caudal vertebrae counts were each significantly different, with C. comus possessing higher counts of each. Snout length, interorbital width, maxilla length, gill-slit length, predorsal length, length of anal-fin connection to caudal fin, and caudal-fin base depth also differed significantly. Among morphometric characters, the following are the most useful to distinguish the species: interorbital width narrower in C. comus (7.0–13.6 % HL vs. 11.9–16.7 % HL in C. faunus ), maxilla length shorter in C. comus (32.0–46.0 % HL vs. 37.8–49.0 % HL), and snout length shorter in C. comus (16.9–28.2 % HL vs. 20.1–34.9 % HL). Differences were also visualized by plotting those morphometric characters most useful for identification against standard length ( Fig. 4 View Fig. 4 ).
The geographic ranges of C. comus and C. faunus overlap in the central and eastern Aleutian Islands. Despite our collection of all liparids encountered in over 1000 tows in the Aleutian Islands during AFSC surveys, only C. comus has been collected west of Adak Island. The two species were obtained syntopically in one haul in Seguam Pass at 291 m depth. This area has become known for its high diversity of liparids, where, for example, three species of Allocareproctus were also collected in the same haul ( Orr and Busby, 2006).
Among other snailfishes of the North Pacific, Temnocora candida ( Fig. 1 View Fig. 1 C) is easily misidentified as either of the new species both when alive and preserved. In life, it is also mottled with red and white but lacks the gray mottling posteriorly of C. comus and C. faunus ( Fig. 1 View Fig. 1 A, 1B). Temnocora candida can also be distinguished from the new species by several other characters, including its having a lobed dorsal fin, lower counts of median-fin rays (D 43–48, A 36–41) and caudal vertebrae (37–42), fewer pyloric caeca (12–13), a wider interorbital width (14.2–26.5 % HL), and a deeper body (22.8–27.3 % at pelvic disk, 19.5– 24.7 % at anal-fin origin). Although T. candida is commonly called the Bigeye Snailfish (Robins et al., 1991; Nelson et al., 2004), both new species have larger orbits (31.6–46.0 % HL in C. comus and C. faunus vs. 21.6–35.2 % HL in T. candida ). The pectoral girdle also differs in having larger proximal radials, with a small fenestra present between proximal radials two and three ( Fig. 2 View Fig. 2 C).
Careproctus bowersianus ( Fig. 1 View Fig. 1 D) was known from two specimens collected on Bower’s Bank in the Bering Sea north of the central Aleutian Islands until Kido (1985) provided a redescription based on additional material from the Aleutian Islands and Bering Sea slope. It is similar in body shape to both new species ( Fig. 1 View Fig. 1 D), and numbers of median-fin rays and vertebrae are within the range of C. comus . However, its overall body coloration, dark stomach, and nonprotruding snout, as well as its smaller mouth, pelvic disk, and orbit immediately distinguish it from each of the new species. The pectoral girdle is also strikingly different, with C. bowersianus having large squarish to rectangular proximal radials that nearly abut one another and lacking fenestrae between proximal radials two and three and between the scapula and proximal radial one ( Fig. 2 View Fig. 2 D).
Among these species, the total number of distal pectoral radials also differs, a character that has not been described for other liparids (Andriashev et al., 1977; Kido, 1988; Andriashev and Stein, 1998). The distal radials in snailfishes are cartilaginous nodules formed in the fibrocartilage pad along the distal margin of the scapulocoracoid cartilage. Typically, the dorsalmost radials are larger, the more ventral radials becoming smaller until they are lost in the one or two ventralmost rays, as in both T. candida and C. bowersianus . In C. comus and C. faunus , the radials become smaller until at rays 22–30 they coalesce with the scapulacoracoid cartilage, upon which the ray bases then articulate. The ventralmost rays articulate freely with the fibrocartilage pad. An analysis of the distribution of this character is beyond the scope of this paper, although it appears worthy of further investigation within the context of liparid phylogenetics.
Three species known only from their holotypes are also similar to C. comus and C. faunus . Careproctus mollis is only known from off Attu Island in the western Aleutian Islands. Fin-ray counts are within the range of C. comus . However, C. mollis is more robust than either new species and has a distinct hump at the nape. It also differs in several morphometric characters, having a greater head width, interorbital width, and caudal-base depth, as well as a smaller orbit.
Careproctus attenuatus was taken in deep waters (882 m) from the western Aleutian Islands off Agattu Island. Its dorsal-, anal-, and pectoral-fin ray counts are at or near the lower end of the range of C. faunus , and it is similar in general body shape to both new species. Unlike the new species, however, its gill cavity is black, its snout does not project beyond the lower jaw, its head is broad and depressed, with the width about equal to the depth ( Gilbert and Burke, 1912), and its dorsal fin is notched slightly ( Burke, 1930; Mecklenburg et al., 2002; not presently evident because of the poor condition of the holotype).
Careproctus sinensis was collected in the Sea of Japan. While its meristics are well within the range of C. comus , its distinctive hump at the nape, similar to that of C. mollis , immediately distinguishes it from C. comus . In addition, the anterior mandibular pores open into a single pit (rather than opening as separate pores in C. comus ) and a single suprabranchial pore is present (two are present in C. comus ).


| N |
Nanjing University |
| W |
Naturhistorisches Museum Wien |
| F |
Field Museum of Natural History, Botany Department |
| V |
Royal British Columbia Museum - Herbarium |
| UW |
University of Washington Fish Collection |
| J |
University of the Witwatersrand |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| CAS |
California Academy of Sciences |
| SIO |
Scripps Institution of Oceanography |
| STAR |
Arkansas State University |
| LACM |
Natural History Museum of Los Angeles County |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |