
Paradialychone, Tovar-Hernández, 2008
|
publication ID |
https://doi.org/ 10.1080/00222930802254714 |
|
persistent identifier |
https://treatment.plazi.org/id/42228538-FFAF-FF9B-FD97-5879FC232102 |
|
treatment provided by |
Felipe |
|
scientific name |
Paradialychone |
| status |
gen. nov. |
Paradialychone View in CoL gen. nov.
Type species
Chone americana Day, 1973 (redescribed by Tovar-Hernández 2005). Diagnosis
Medium to large-bodied sabellin species. Branchial lobes fused dorsally. Radiolar skeleton with two rows of cells. Palmate membrane and radiolar flanges present. Dorsal lips elongate, longer than wide, without branchial skeleton extensions. Ventral lips present, broadly rounded. Dorsal pinnular appendages and ventral radiolar appendages present. Pinnules of equal-size along the entire radiole. Anterior margin of anterior peristomial ring triangular or bilobed. Posterior peristomial ring collar present. Glandular ridge on chaetiger 2 narrow or broad laterally. Ventral shields absent. Notopodia in chaetiger 1 with two groups of elongate, narrowly hooded chaetae in C-shaped arrangement. Notopodia in chaetigers 2–8 with superior group of chaetae in two irregular rows, all elongate, narrowly hooded; inferior group with one anterior row of short bayonet chaetae, two posterior rows with symmetrical, mucronate paleate chaetae. Thoracic neuropodia bearing acicular uncini with a large tooth offset from midline, followed by series of smaller teeth; dentition covering three-quarters of the main fang length; hood present, handles long. Anterior abdominal segments with two transverse rows of elongate, narrowly hooded chaetae, chaetae from the anterior row shorter than chaetae in posterior row; uncini with well-developed rectangular breast, handles absent, main fang well developed with a large tooth above the main fang in midline, followed by a series of smaller teeth covering one-half of the main fang length; intrafascicular variation: older uncini, located dorsalmost in torus, smallest in size, younger uncini, located ventralmost in torus, biggest in size. Posterior abdominal segments with very elongate, narrowly hooded chaetae; modified uncini with hooked breast, handles absent and dentition covering three-quarters of the main fang length; intrafascicular uncinal variation similar to those in anterior abdominal segments. Pre-pygidial depression simple.
Etymology
The genus name is made by combining the Greek word Para, meaning beside or near and the genus name Dialychone to emphasize their morphological resemblance.
Remarks
Paradialychone gen. nov. is monophyletic on the basis of anterior abdominal uncini with a large tooth above the main fang, located in midline, followed by a series of smaller teeth. Dialychone and Paradialychone gen. nov., are sister groups sharing the presence of elongate dorsal lips, lacking branchial skeleton extensions [3(2)] and a simple pre-pygidial depression [52(1)], in contrast to Chone , in which dorsal lips are broadly rounded and lack pre-pygidial depression; or in Euchone , where the dorsal lips are elongated but have radiolar appendages or mid-rib and the pre-pygidial depression is often provided with lateral wings ( E. analis ) or with an anterior margin ( E. pseudolimnicola ). In previous studies, Euchone was diagnosed by the presence of a pre-pydigidial depression ( Cochrane 2003; Giangrande and Licciano 2006); as shown above, its use as a diagnostic feature for an independent genus must be discontinued because Dialychone and Paradialychone gen. nov. share this feature.
Paradialychone gen. nov. differs from Dialychone by having abdominal uncini with a large tooth above the main fang, located in midline, followed by a series of smaller teeth (with a series of teeth of nearly uniform size in Dialychone ). The genus contains 12 newly combined species: Paradialychone americana n. comb.; P. bimaculata n. comb.; P. diazi n. comb.; P. ecaudata n. comb.; P. eiffelturris n. comb.; P. farringtonae n. comb.; P. filicaudata n. comb.; P. gambiae n. comb.; P. johnstonae n. comb.; P. paracincta n. comb.; P. paramollis n. comb.; and P. uebelackerae n. comb., and one species not named formally and referred as Chone sp. British Virgin Islands by Tovar-Hernández (2007b).
Concluding remarks
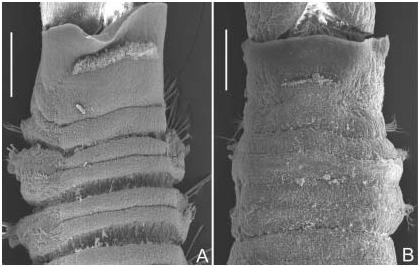
Chone View in CoL , Dialychone View in CoL and Paradialychone View in CoL gen. nov., are monophyletic genera, but the strict consensus tree shows some polytomies inside them. Nevertheless, their resolution would be improved once new morphological characters are found and employed, particularly in addition to the presence or absence of the branchial skeleton extension of dorsal lips based on detailed histological techniques. Also, the presence or extension of the ventral patch of cilia is an interesting property that has not been detailed before, thus, in P. ecaudata View in CoL gen. nov. ( Figure 9A View Figure 9 ) and D. trilineata View in CoL n. comb., ( Figure 9B View Figure 9 ) the ventral shield of the collar is completely ciliated, but the cilia are not an extension of those present in the anterior peristomial ring, as occurs in some members of Laonome Malmgren, 1866 View in CoL (Fitzhugh, personal communication) or even in Amphicorina View in CoL . Most species within Chone View in CoL sensu stricto have a large, swollen ventral collar shield, but in other such as D. acustica View in CoL , it is vestigial or poorly developed. Therefore, the detailed observation of these structures using scanning electronic microscopy could add new features to the study of chonids.
Another interesting feature that deserves further consideration from both taxonomic and systematic standpoints is the relative development of the glandular ridge epithelium because externally and internally, the glandular ridge varies among species of Chone View in CoL as emphasized by Tovar-Hernández and Sosa-Rodríguez (2006). For example, in species with a narrow glandular ridge, such as C. infundibuliformis View in CoL , P. ecaudata View in CoL gen. nov., C. mollis View in CoL and D. quebecensis View in CoL n. comb., it occupies the external half of the glandular columnar epithelium ventrally whereas dorsally, the glandular ridge occupies only a very small area. In others species, as in D. veleronis View in CoL n. comb., it occupies three-quarters of the epithelium. Furthermore, the relative size and distribution of oocytes, and the spermatozoal morphology are probably the most important features needing additional clarification. Some species have hundreds of small oocytes distributed in the thorax and abdomen, whereas in others, a few large oocytes are distributed on thoracic and abdominal segments ( Tovar-Hernández 2007a). Unfortunately, this information is only available for a few species, summarized in the series of papers on the revision of Chone View in CoL ( Tovar-Hernández 2005, 2007a, 2007b; Tovar-Hernández and Sosa-Rodríguez 2006; Tovar-Hernández et al. 2007).
The previous series of papers dealing with a worldwide revision of Chone species together with this cladistic analysis represent a complete effort to fully understand a group of plesiomorphic sabellids. However, the genera Amphicorina , Euchone and Jasmineira remain confusing. The species currently recognized within Amphicorina have noticeable variations in chaetal morphology: (1) superior thoracic notochaetae could be elongate, narrowly hooded or paleate; (2) inferior thoracic notochaetae could be short bayonet chaetae, or short capillaries; and (3) abdominal neurochaetae needle-like, or elongate, broadly hooded. On the other hand, Cochrane (2003) and Giangrande and Licciano (2006) pointed out the high variability of the species recognized within the paraphyletic genus Euchone , promoting the examination of the species. Finally, Jasmineira also displays variations that merit further clarification (except for J. candelus Grube, 1863 , J. oculatus Langerhans, 1884 , now recognized within Clavirramus Fitzhugh, 2002), for example, in some species the posterior row of inferior thoracic notochaetae is composed by paleate chaetae, and for others these are represented by short, broadly hooded chaetae. Thus, Amphicorina , Euchone and Jasmineira are genera worthy of worldwide revisions; the examination based on typematerial coupled with phylogenetic analyses could solve their taxonomy.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Paradialychone
| Tovar-Hernández, María Ana 2008 |
Paradialychone
| Tovar-Hernández 2008 |
P. ecaudata
| Tovar-Hernández 2008 |
D. trilineata
| Tovar-Hernández 2008 |
P. ecaudata
| Tovar-Hernández 2008 |
D. quebecensis
| Tovar-Hernández 2008 |
D. veleronis
| Tovar-Hernández 2008 |
C. mollis
| Bush 1904 |
Laonome
| Malmgren 1866 |
Chone
| Kroyer 1856 |
Chone
| Kroyer 1856 |
Chone
| Kroyer 1856 |
C. infundibuliformis
| Kroyer 1856 |
Chone
| Kroyer 1856 |