
Hypostomus chrysostiktos, Birindelli & Zanata & Lima, 2007
|
publication ID |
https://doi.org/10.1590/S1679-62252007000300006 |
|
DOI |
https://doi.org/10.5281/zenodo.17101303 |
|
persistent identifier |
https://treatment.plazi.org/id/4116BF05-FFA2-F604-F392-FDE1FA8F1244 |
|
treatment provided by |
Carolina |
|
scientific name |
Hypostomus chrysostiktos |
| status |
sp. nov. |
Hypostomus chrysostiktos View in CoL , new species
Figs. 1 View Fig and 2 View Fig
Pterygoplichthys sp. : Higuchi, Britski & Garavello, 1990: 225 [misidentification; Bahia, rio Paraguaçu at the area currently under Pedra do Cavalo dam].
Holotype. MZUSP 88157 , 222.4 mm SL, Brasil, Bahia, Iaçu, rio Paraguaçu , Fazenda Santo Antonio at 10 km from Vila São Vicente, 12°30’54.1”S 39°22’53.9”W, 134 m elevation, 7 Jun 2005; A. M. Zanata, J. L. O. Birindelli, O. T. Oyakawa, M. P. Geraldes, P. C. A. Cardoso & P. Moura. GoogleMaps
Paratypes. All from Brazil, Bahia, rio Paraguaçu basin: ANSP 185374 About ANSP , 1, 166.6 mm SL; GoogleMaps MZUSP 88158 View Materials , 3, 157.9- 212.2 mm SL; collected with holotype. GoogleMaps AUM 45646 , 1, 117.0 mm SL; GoogleMaps MZUSP 88159 View Materials , 5, 72.7-223.5 mm SL; GoogleMaps UFBA 02786 , 5, 112.3- 259.7 mm SL; Iaçu, rio Paraguaçu , Fazenda Os Touros , 12°41’10.6”S 40°07’4.7”W, 143 m elevation, 8 Jun 2005; A. M. Zanata et al.. GoogleMaps MCP 41000 , 1, 101.0 mm SL; GoogleMaps MZUSP 91635 View Materials , 3, 96.5-149.5 mm SL; Itaetê, rio Una , 12°56’08”S 41°03’54”, 4 Dez 2005; A. M. Zanata et al. GoogleMaps MNRJ 22780 View Materials , 2, 50.4- 147.3 mm SL; Palmeiras, rio Santo Antônio ( Bahema ), 12°25’16.6”S 41°22’20.9”W, 8 Nov 1999; A. Clistenes. GoogleMaps MNRJ 22781 View Materials , 1, 151.6 mm SL; Palmeiras, rio Santo Antônio ( Camelo ), 12°25’36.7”S 41°25’49.1” W, 8 Nov 1999; A. Clistenes. GoogleMaps MZUSP 49262 View Materials , 1, 45.5 mm SL; GoogleMaps MZUSP 84231 View Materials , 3 (1 c&s), 29.0- 33.1 mm SL; Itaetê, Olho d´água do Almerindo , tributary of rio Una , Jun 1993; A. M. Zanata et al. GoogleMaps MZUSP 88161 View Materials , 2, 68.4-104.5 mm SL; GoogleMaps UFBA 02788 , 2, 79.4-93.4 mm SL; rio Ferro Doido at Cachoeira Domingos Lopez , rio Jacuípe basin, 11°33’32.1”S 40°54’20.3”W, 662 m elevation, 11 Jun 2005; A. M. Zanata et al. GoogleMaps MZUSP 88165 View Materials , 2 (1 c&s), 117.5-122.7 mm SL; GoogleMaps MZUSP 88617 , 2 (1 c&s), 114.0- 194.8 mm SL; GoogleMaps UFBA 02031 , 3, 60.3-82.3 mm SL; GoogleMaps UFBA 02040 , 23, 78.2-163.8 mm SL; GoogleMaps UFBA 02785 , 6, 129.4- 222.8 mm SL; Cachoeira, rio Paraguaçu (area currently under Pedra do Cavalo dam ), 15 Oct 1980; V. Almeida et al.. GoogleMaps MZUSP 88173 View Materials , 1, 117.9 mm SL; GoogleMaps UFBA 02039 , 2, 115.2- 133.4 mm SL; Lençóis, rio Santo Antônio de Licurioba , 5 Feb 2001; L. Panelli. GoogleMaps UFBA 02030 , 2, 100.4- 153.9 mm SL; Cachoeira, rio Paraguaçu (area currently under Pedra do Cavalo dam ), 13Aug 1980; V.Almeida et al. GoogleMaps UFBA 02787 , 3, 211.9- 223.9 mm SL; Iaçu, rio Paraguaçu , Fazenda Os Touros , 12°41’10.6”S 40°07’4.7”W, 143 m elevation, 9 Jun 2005; A. M. Zanata et al GoogleMaps .
Diagnosis. Hypostomus chrysostiktos is readily distinguished from most loricariids, with the exception of Megalancistrus , a few species of Chaetostoma , and some species of Pterygoplichthys and Delturus , by the high number of dorsal-fin branched rays (10-11). It can also be distinguished by having slightly evertible cheek plates ( vs. fully evertible cheek plates of Chaetostoma , Megalancistrus , and most other Ancistrini and Pterygoplichthyini taxa; or no evertible cheek plates in Delturinae , Neoplecostominae, Hypoptopomatinae , Loricariinae , Corymbophanini and Rhinelepini). It can be further distinguished from Delturus by having four branched anal-fin rays ( vs. five or six branched anal-fin rays). In addition, the new species differs from several loricariids by having naked abdomen ( vs. plated in e.g. Pterygoplichthys ), snout almost completely plated leaving just a small naked area on its tip ( vs. snout completely naked in e.g. Chaetostoma ). See Discussion for additional notes on its diagnosis.
Description. Standard length of examined specimens 29.0 to 259.7 mm. Counts and measurements of holotype and paratypes in Tables 1 View Table 1 and 2 View Table 2 . Dorsal, lateral, and ventral views of head and body in Figure 1 View Fig . Dorsal profile of body convex from snout tip to dorsal-fin origin, and straight from this point to middle of caudal peduncle. Ventral profile of body straight from snout to caudal-fin base. Body relatively depressed; greatest body width at cleithral region, progressively tapering posteriorly from cleithrum. Greatest body depth at dorsal-fin origin, gradually tapering to caudal peduncle. Caudal peduncle robust and roughly trapezoid in cross-section; somewhat flattened on ventral portion.
Head somewhat depressed, wide, and round anteriorly. Head without crests. Posterior border of supraoccipital straight to somewhat irregular in form, without distinct posteriorly directed expansion. One plate bordering posterior margin of supraoccipital bone. Interorbital region straight, with lateral margins elevated. Eyes relatively small. Dorsal flap of iris present. Mouth distinctly wide; roof with relatively small median buccal papilla. Lips rounded, of moderate size, occupying approximately two-thirds of ventral surface of head, with fringed edges. Inner surfaces of lips covered by papillae, except for smooth region just behind toothed portions of dentaries; smooth region wider posterior to symphysial region. Anteriormost papillae of inner surface of lower lip roundish and relatively large, followed by patch of smaller and closely positioned papillae; anteriormost papillae in inner surface of upper lip roundish and small, followed by patch of larger and more elongated papillae. Teeth long and bicuspid; larger cusp approximately 1.5 to 2.0 times length of outer cusp and curved inward, similar to Hypostomus auroguttatus ( Muller & Weber, 1992: fig. 2e). Contralateral premaxillary teeth forming relatively straight line or slightly concave arch; contralateral dentaries forming relatively wide angle and dentary teeth forming concave arch facing mouth cavity. Maxillary barbel short, usually shorter than orbital diameter.
Dorsal-fin origin situated on vertical anterior to pelvic-fin origin and approximately on midlenght of pectoral-fin spine. Tips of first and last basal radials of dorsal fin lying above neural spines of vertebrae seven and 19, respectively. Dorsal fin large; tips of adpressed last rays almost reaching to slightly surpassing posterior border of preadipose plate. Margin of dorsal fin slightly convex. Adipose-fin spine narrow, usually straight or slightly curved ventrally. Pectoral fin of moderate size with posterior margin straight to slightly concave. Degree of overlaping of posterior tip of adpressed pectoral fin over pelvic fin ontogenetically variable; tip of pectoral reaching middle of pelvic fins in smaller and median-sized specimens, and reaching posterior third of pelvic-fin spine in largest individuals. Pectoral fins inserted about same plane as pelvic fins and pectoral-fin spine resting on top of pelvic-fin when adpressed. Pelvic fin of moderate size, with posterior margins rounded in smaller and straight in larger specimens. Tip of pelvic fin reaching origin of anal-fin rays. Plate of first proximal radial of anal fin covered by skin in all specimens examined. Tips of first and last basal radials of anal fin lying below hemal spines of vertebrae 16-18, respectively. Caudalfin margin concave, lower spine longer than upper. Thirty vertebrae, four branchiostegal rays.
Body entirely covered by plates, except for ventral surface from mouth to anus and areas around dorsal, pectoral, pelvic, and anal-fin insertions. Cheeks, snout, and rostrum completely plated, except for naked area on snout tip that continues ventrally to margin of upper lip, and small areas around nares. Ornamentation of pterotic-supracleithrum usually similar to remaining surface of head and with odontodes densely distributed, arranged in well-defined rows, diverging posteriorly in larger specimens. Plates over opercle weakly armed, varying from covered with relatively small odontodes similar in size with those in nearby areas to posterior portion of opercle with slightly more elongated odontodes (up to 22). Posterior portion of preopercle surface varying from having small odontodes to up to twenty slightly elongate odontodes. Cheek plates slightly evertible. Median series of plates bearing lateral-line canal. Dorsal series of plates starting posteriorly, at vertical through dorsal-fin origin. Ventral series of plates usually starting at midlength of pelvic-fin base. Dorsalmost plates between end of dorsal-fin base and adipose-fin spine flattened. One preadipose plate. Five rows of plates on the caudal peduncle.
Plates on dorsal and lateral portion of body relatively smooth in all specimens examined. Defined keels present only in mid-ventral series of plates, more developed on first five plates. Weakly developed crest in mid-dorsal series of plates of some individuals. Plates situated on dorsal and lateral portion of body covered with minute odontodes, clearly organized in rows diverging posteriorly; odontodes usually more developed on borders of plates of mid-dorsal and midventral series. All fin rays covered by small odontodes, more conspicuous in dorsal-fin rays and adipose spine. Dorsal and lateral surfaces of anterior portion of pectoral fin spine covered with relatively small odontodes, increasing in size and curvature towards posterior portion of pectoral-fin spine; odontodes more developed in specimens of 180.0 mm SL or larger. Dorsal surface of branched pectoral-fin rays with small odontodes along their entire lengths; odontodes more conspicuous on larger individuals. Pelvic-fin spine and dorsal surface of all branched rays covered with small odontodes along their entire length. Caudal-fin rays covered with odontodes, more evident on unbranched rays.
Color in alcohol. Overall ground color of dorsal and lateral surface of body brownish. Body covered with small roundish clear spots. Light brown spots on head and dorsal portion of trunk. Spots usually more evident and larger on lower lateral portion of body. Spots on dorsal portion of head relatively smaller than those of trunk, less interspaced and decreasing in size towards snout. Trunk and fins covered with comparatively larger spots; some specimens around 90 mm SL with longitudinally-aligned spots. Naked ventral surface of head and body (including lips) pale to yellowish from area behind lower lip to area close to anal-fin insertion. Naked area of snout and upper lip covered with small clear spots (more evident on UFBA 02788 specimens). Ventrolateral margins of head and body with dark coloration and large pale spots and vermiculations at boundaries between plated and naked areas in most specimens examined; specimens around 100 mm SL with dark vermiculations across coracoid region. Ventral portion of caudal peduncle usually dark brown, with clear spots and/or vermiculations sparsely distributed.
Overall ground coloration of all fins dark brown with spots scattered distributed over spines, rays and inter-radials membranes. Spots over fins usually similar in size to those distributed over trunk. Caudal-fin spots usually paler than those of remaining fins. Spots of paired fins clearer from ventral view. Some specimens, often those with very dark overall pigmentation, with spotted pattern described above less visible; spots nearly completely absent in some individuals.
Color in life. Description based on field observations of several specimens by JLOB and AMZ, and on pictures of living specimens (see Fig. 2 View Fig ). Ground color brown with bright yellow spots, much more evident than in preserved specimens. Ventral surface of body yellowish. Yellow spots on fins more conspicuous over rays.
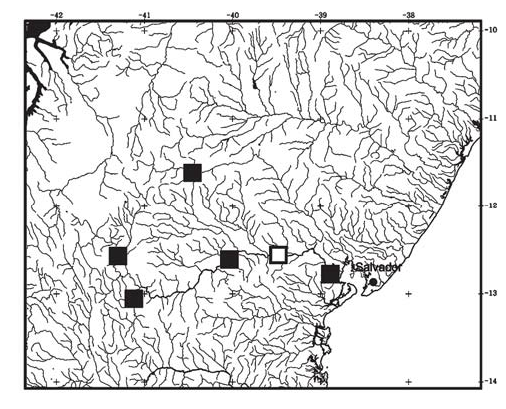
Distribution. Only known from rio Paraguaçu basin, an independent coastal drainage in northeastern Brazil, where it is widespread ( Fig. 3 View Fig ).
Etymology. From the greek chrysos, after gold, and stiktus, after spotted, in allusion to the bright yellow spots present in living specimens. An adjective.
Ecological notes. Hypostomus chrysostiktos was collected in dark water stretches of the rio Paraguaçu and its tributaries, with rocky bottom, at altitudes ranging from 50 to 662 meters above sea level.
Phylogenetic relationships. The 215 characters previously defined by Armbruster (2004) for loricariids and outgroups were coded for H. chrysostiktos ( Table 3 View Table 3 ). Hypostomus chrysostiktos was then added to Armbruster’s (2004) matrix. The resulting phylogenetic hypothesis recovered H. chrysostiktos as the sister group of a clade composed by H. emarginatus group plus H. commersoni and, consequently, within the Hypostomini tribe (sensu Armbruster, 2004). Armbruster (2004: 51) found two well-corroborated monophyletic entities within the tribe Hypostomini , the H. emarginatus group (including Aphanotolurus and Isorineloricaria ) and the H. cochliodon group. Nevertheless, “there are no general trends in the relationships of the remainder of the species” ( Armbruster, 2004: 51). Hypostomus chrysostiktos does not have the diagnostic characters for any of the two afore-mentioned groups (see Armbruster, 2004: 51).
Hypostomus chrysostiktos is included in the subfamily Hypostominae by possessing the following characters: a long acessory process on the first ceratohyal (character 7: state 2 of Armbruster, 2004), the lower lobe of hypural plate longer than the upper (123: 1), a V-shaped spinelet (148: 0), and a posteroventral ridge on the basipterygium (173: 1). Hypostomus chrysostiktos belongs to the clade including Hypostomini , Pterygoplichthyni, and Ancistrini by the presence of eight derived characters ( e.g., anterior process of pterotic-supracleitrum separated mesially from main body, connected by a strut (111: 1); four branched anal-fin rays (150: 2), and slightly evertible cheek plates (184: 1). Finally, H. chrysostiktos shares with the members of the currently defined Hypostomini the presence of a hatched-shape opercle (78:1), the anterior process of the pterotic-supracleitrum passing halfway through the orbit (112: 1), and a pointed post-cleithral process (156: 1).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |