
Napaeus teobaldoi Martín, 2009
|
publication ID |
https://doi.org/ 10.1080/00222930903094621 |
|
persistent identifier |
https://treatment.plazi.org/id/4014878B-456E-7939-4092-8436FD09FC2B |
|
treatment provided by |
Felipe |
|
scientific name |
Napaeus teobaldoi Martín |
| status |
sp. nov. |
Napaeus teobaldoi Martín View in CoL sp. nov.
Type material
Holotype ( Figures 4S View Figure 4 , 7D View Figure 7 ): TFMC ( MT 0399 ). Type locality: Barranco Hondo , in the southeastern slope of La Culata mountain (Tenerife; UTM coordinates: 28 RCS6742 View Materials ; 270 m altitude). Leg. Javier Martín, 7 May 2008
Paratypes: two specimens and four shells, collected in 2008 from the type locality and deposited in CML (four shells) and AIT .
Etymology
The specific name derives from Teobaldo, the name of Javier Martín’s father, to whom this species is dedicated.
Geographic range and habitat
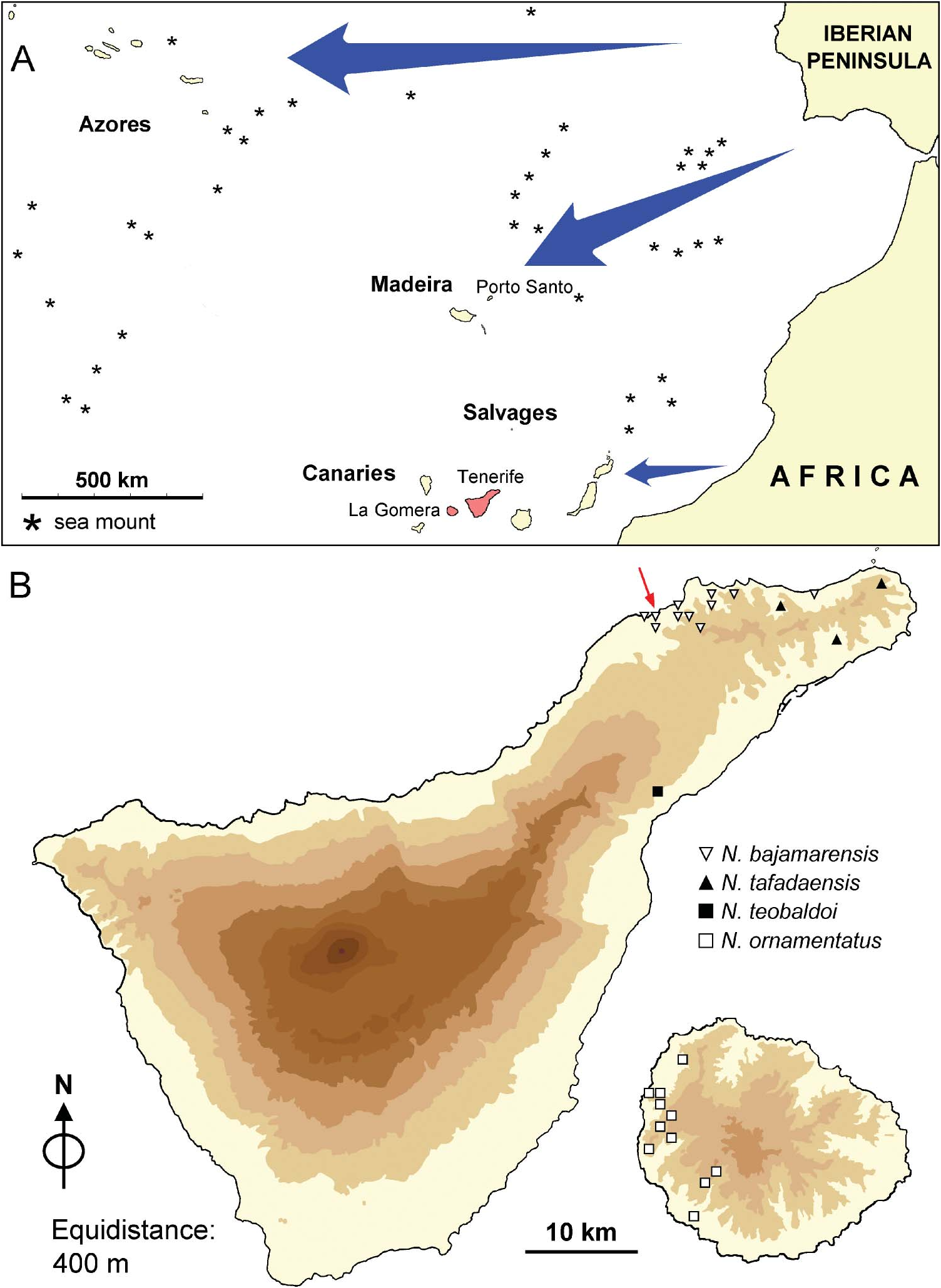
Endemic to Tenerife, only present in the type locality, associated with lowland vegetation ( Figure 1B View Figure 1 ). It is a ground dweller; living specimens were found buried to some cm deep.
Diagnosis
Shell elongated, cone-shaped, with a very regular growth, uniform brown coloured. Teleoconch with very numerous radial oblique ribs, regularly spaced in the first whorls and frequently interrupted, irregularly undulating, giving the appearance of granulation in the rest. Epiphallus with an epiphallar caecum. Bursa copulatrix duct without diverticulum.
Description
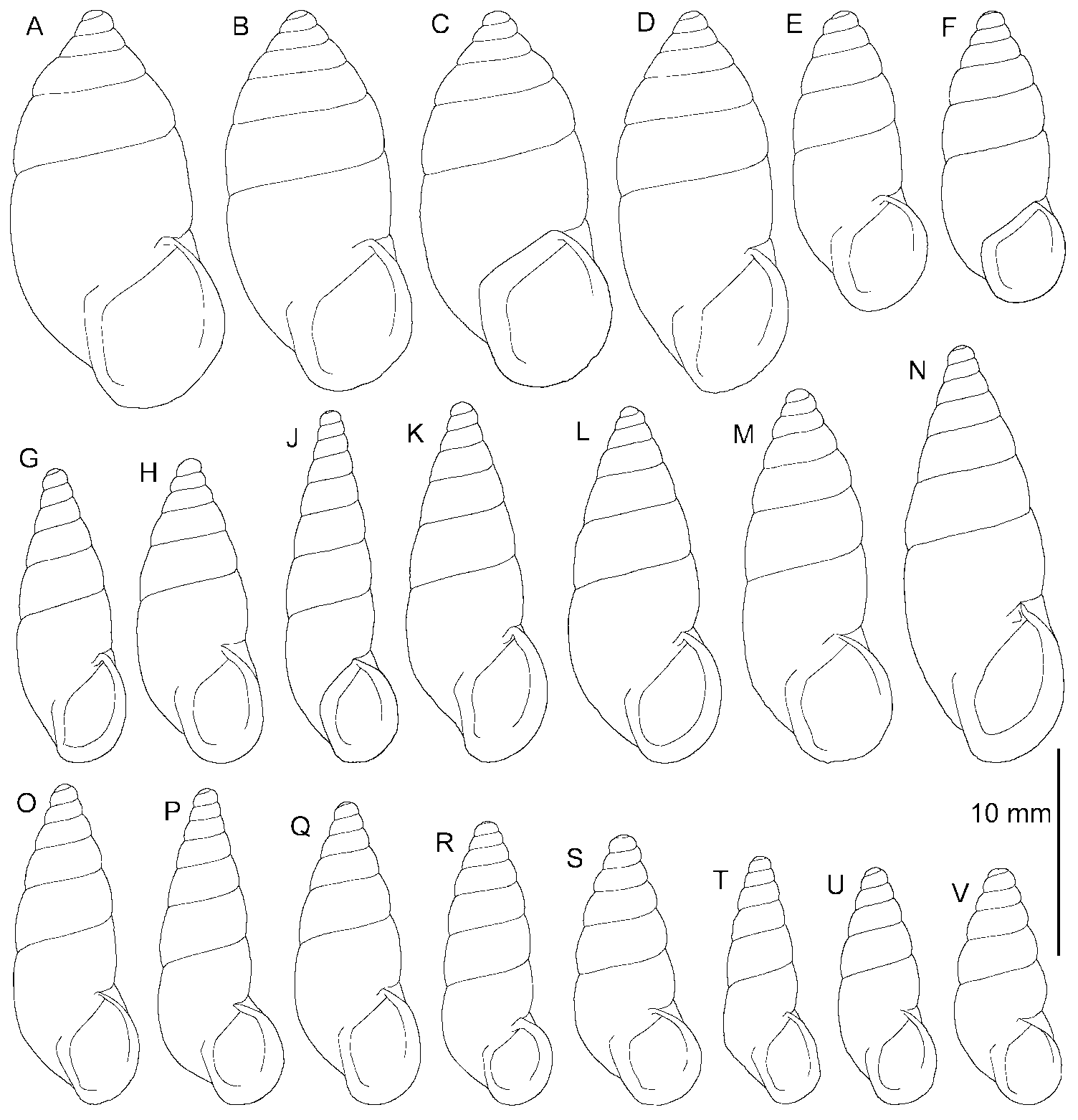
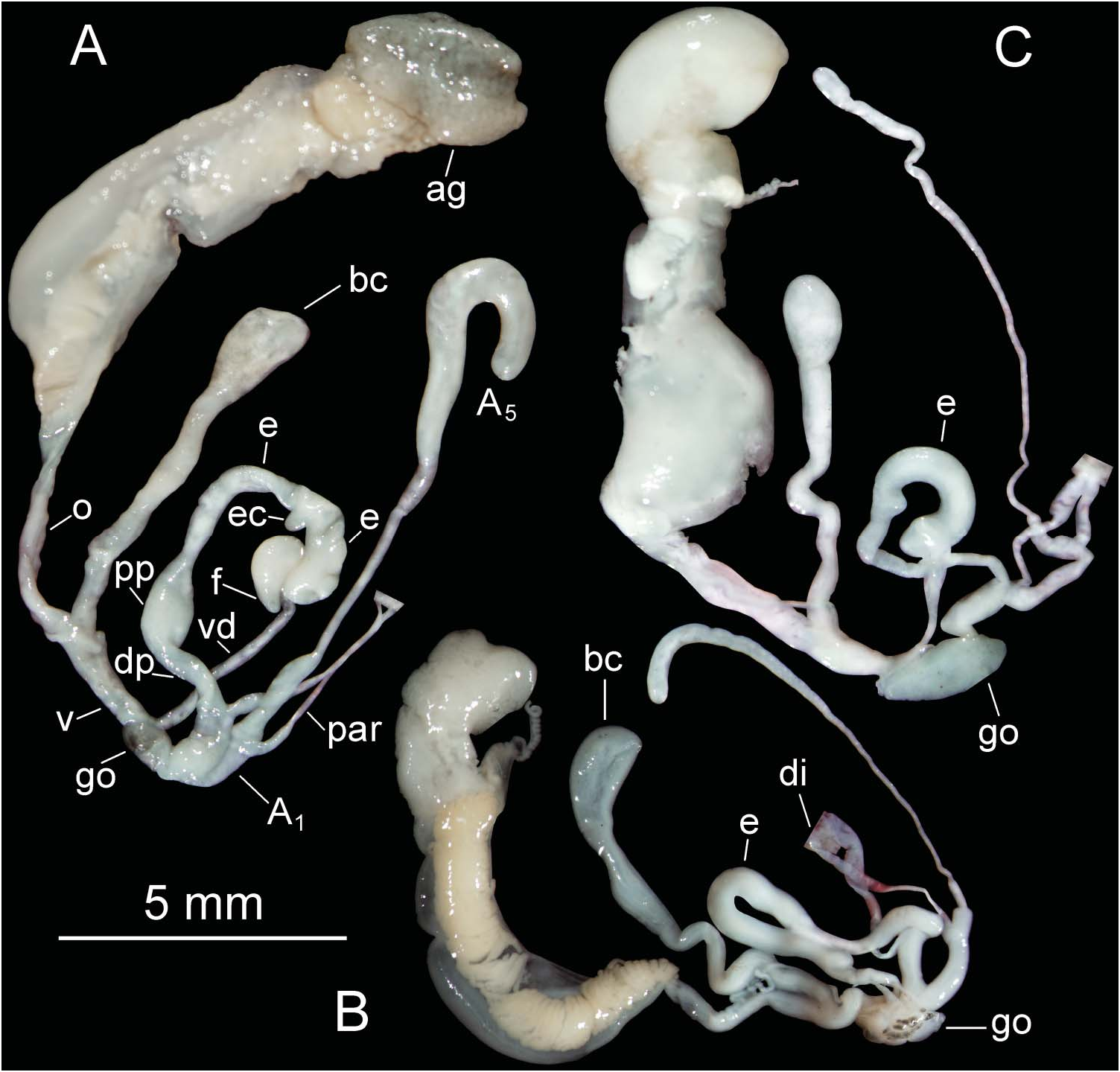
Shell ( Figure 7D View Figure 7 ) obese (SB/SH index), elongated cone-shaped, about 6¾–7 convex whorls, deeply marked suture. Body whorl intermediate (BH/SH index) and protoconch smooth, with two whorls. Aperture short, wide (AH/SH and AB/SB indices, respectively), elliptical section at the palatal side, slightly angular at union of columellar and palatal edges. Upper palatal side forms angle greater than 90º with parietal side, aperture juts out considerably from body whorl beginning. Whitish discontinuous peristome expanded as lip, more developed in lower part of palatal edge and reflected in columellar edge, where it partially covers umbilical slit. Older specimens display callosity between peristome edges, nodule at union of parietal–palatal area.
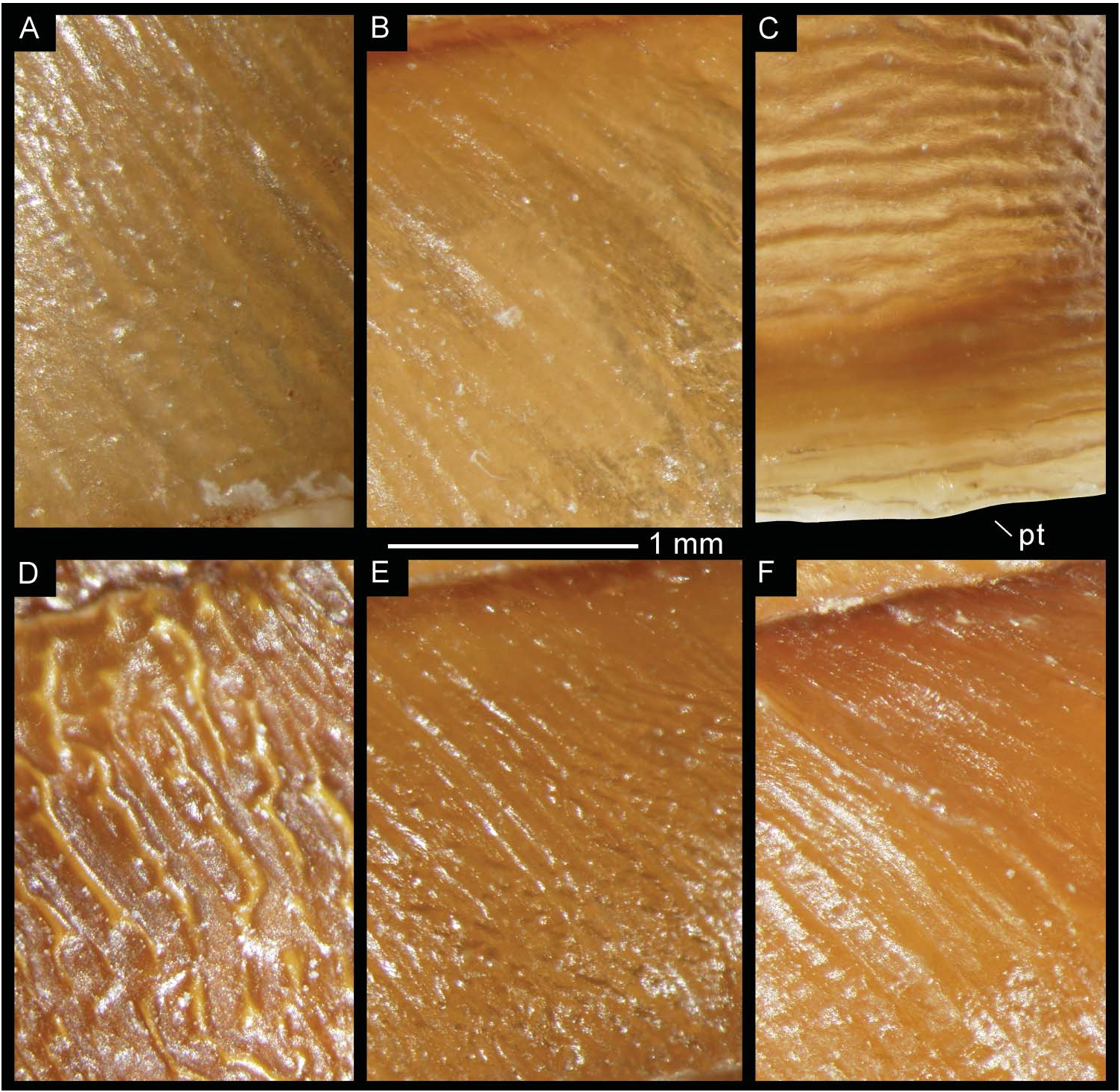
Shell uniform brown coloured. Ornamentation characterized by very numerous radial oblique ribs, regularly disposed in first whorls. From third/fourth whorls, some shell ribs sinuous, many shell ribs interrupted, irregularly undulating ( Figures 7D View Figure 7 , 10E View Figure 10 ) giving appearance of granulation. At columellar zone of body whorl, all ribs sinuous, irregularly undulating.
Genital system ( Figure 11 C View Figure 11 ). Genital system not fully developed in three live specimens collected (holotype and two paratypes). Atrium very short. Penis and epiphallus tubular, without clear delimitation between both in three specimens. Only distal penis portion clearly distinguishable, located between atrium and insertion of penial retractor muscle. Epiphallus with epiphallar caecum and widened proximal region. Flagellum very short. Vas deferens opens laterally on proximal epiphallar end. Penial appendix about twice length of penis and epiphallus together, arising near proximal end of distal penis portion. A 1 appendix penial portion about twice length of distal penis portion. Appendicular retractor muscle inserts very close to penial retractor on diaphragm walls and, laterally, about midpoint of A 1 portion. Free oviduct longer than vagina. Bursa copulatrix complex without diverticulum.
Remarks
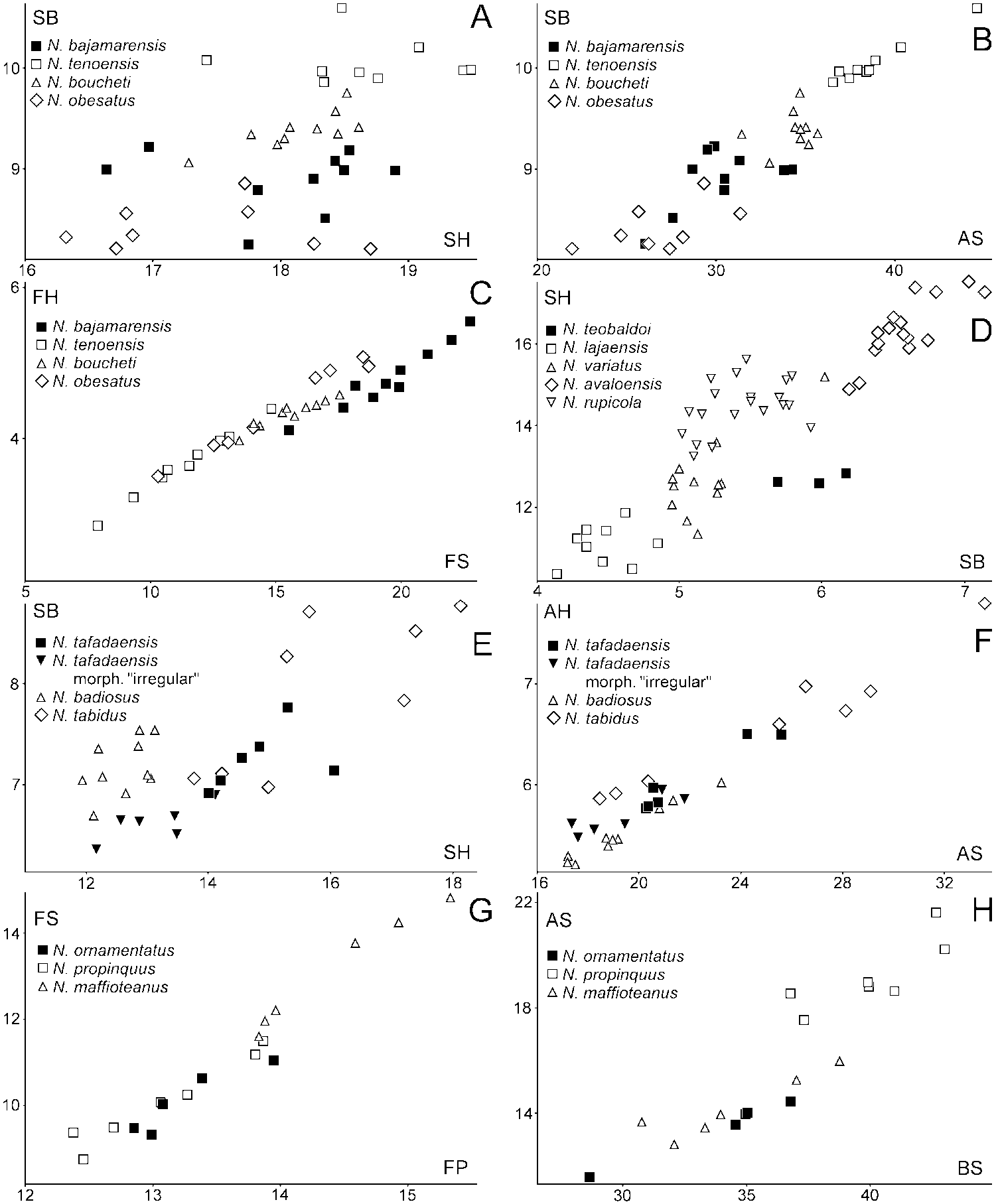
Shell morphology. The shell of N. teobaldoi sp. nov. ( Figure 4S View Figure 4 ) has very regular growth, being cone-shaped. Few other Napaeus species have such regular growth. These include N. variatus ( Figure 4P View Figure 4 ), N. rupicola ( Figure 4J View Figure 4 ), N. avaloensis ( Figure 4K View Figure 4 ) and the Pleistocene extinct N. lajaensis ( Figure 4T View Figure 4 ), but they are more slender and their dimensions differ from N. teobaldoi ( Figure 6D View Figure 6 ).
Genital system anatomy. The genital system of N. teobaldoi has an epiphallar caecum which is lacking in N. variatus , N. rupicola and N. avaloensis .
Taxonomic complexity and relationships of Napaeus
The Canarian enid species
Study of the genital system of N. baeticatus , N. tenoensis , N. boucheti ( Henríquez et al. 1993a,b) and N. propinquus (unpublished data), together with that of N. tafadaensis ( Figure 11A View Figure 11 ) and N. teobaldoi ( Figure 11C View Figure 11 ) described here, reveals contradictions with the original Hesse (1933) subgenera descriptions. These six species share two important character-states of both Hesse subgenera: the presence/absence of a diverticulum in the bursa copulatrix duct and/or an epiphallar caecum. The absence of a diverticulum is the main character-state of Napaeus (Napaeus) and the presence of an epiphallar caecum is a character-state of Napaeus (Napaeinus) . The six species cited earlier lack a diverticulum in the bursa copulatrix duct as in Napaeus (Napaeus) , but possess an epiphallar caecum as in Napaeus (Napaeinus) . These dissimilarities were also highlighted by Alonso, Goodacre, et al. (2006), who correlated anatomical and molecular data in only one of the characters used by Hesse (1933) in the original description of the Napaeus subgenera, the character-states “epiphallus with/without a caecum”. As Bank and Neubert (1998) state, the systematic significance of the presence or absence of a diverticulum is probably irrelevant and its presence or absence does not reflect natural affinities. Thus, the new species are not assigned to the Hesse Napaeus subgenera until a phylogenetic analysis of the genus is conducted.
Moreover, Henríquez et al. (1993b) showed two different patterns of spermatophore: with/without a spur-like hook. They also found a positive correlation between the presence of the epiphallar caecum and the spur-like hook of the spermatophore, a relationship confirmed here with the new species N. bajamarensis . Several spermatophoral architectures have been described previously ( Henríquez et al. 1993b; Alonso et al. 1995). These architectures relate to the internal anatomy of the epiphallus ( Ibáñez et al. 2007) and the flagellum, a fact which validates the utility of these characters within taxa classification and description. Therefore, the study of the spermatophore appears to be a useful, complementary method for understanding the taxonomic relationships among Napaeus species.
The Macaronesian enids
The relationships among Macaronesian enids are unknown due to the absence of phylogenetic analyses. However, it is possible to hypothesize potential relationships based on the available taxonomic data.
There is a clear, anatomical difference between Napaeus and Macaronapaeus based on the penial appendix. The presence of the penial appendix is a symplesiomorphy of the Canarian Napaeus shared by several other enids. This character was used by Forcart (1940) for the division of the Enidae into two subfamilies, Eninae (with a penial appendix) and Chondrulinae (without it). However, Bank and Neubert (1998), utilizing the data of Gittenberger (1978, 1983) and Bank (1985), pointed out that the penial appendix has been independently lost in at least six Enidae groups, including the group containing Macaronapaeus . Bank and Neubert (1998) considered the treatment of Napaeus and Macaronapaeus as genera or subgenera a matter of opinion and subsequently placed Macaronapaeus as a subgenus of Napaeus . However, the absence of enids in the Madeira archipelago together with the respective absence/presence of the penial appendix in the Azorean/Canarian enids and the large distance separating those two populations suggest not only a different immigration pathways ( Figure 1A View Figure 1 : arrows) but also potentially different phylogenetic origins.
Several authors have noted a lack of homogeneity within and between the different plant communities of the Macaronesian archipelagos (Azores, Madeira, Salvages, Canary Islands and Cape Verde) and suggested that the observed heterogeneity may be the result of independent colonizations events from different mainland sources ( Beyhl et al. 1995; Borgen 1995). Waldén (1984) indicated that “the most conspicuous difference between the mollusc faunas of Madeira and the Canary Islands is the complete absence of taxa with northwest African affinities on Madeira, despite the fact that they hold a similar position relative to the African continent. “Porto Santo, the oldest island of the Madeiran archipelago, was susceptible to colonization for a period of 10 million years without major volcanic activity. Colonizers arrived by island-hopping from Europe (specifically, from the southwest Iberian Peninsula) through the chain now remaining as sea mounts ( Cook 2008). The western and northernmost Azorean islands – its oldest island, Santa Maria, is estimated to have a maximum age of 8.12 million years, with S. Miguel following with an age of 4.01 million years (Borges et al. forthcoming 2009) – were probably colonized directly from Europe. By this mode of expansion, Arnedo et al. (2001) showed that the spider dysderid species of the Azores Islands are certainly the result of independent colonization from Europe, unrelated to those of the Madeiran and Canarian archipelagos.
Alonso et al. (2000) and Arnedo et al. (2001) suggested that the Canary Islands were colonized from neighbouring northwest Africa. This hypothesis is also supported by the presence of several species of the genera Obelus Hartmann, 1842 ( Ibáñez et al. 2003) and Theba Risso, 1826 ( Gittenberger and Ripken 1985, 1987; Gittenberger et al. 1992) in both regions.
The eastern Canary Islands (Lanzarote and Fuerteventura) are the oldest islands (15–20 million years; Carracedo et al. 2005) only about 115 km distant from the northwest African coast. Sea level fluctuations during recent glacial events decreased that distance to as little as ∼ 70 km ( García-Talavera 1997, 1999). During the Miocene–Pliocene, northwest Africa was wetter, supporting abundant vegetation (see recent review in Swezey (2009)) and mighty river systems including the Drâa, which is located opposite Lanzarote and Fuerteventura. The Drâa currently has a wide valley (4–5 km) and a relatively short course, both of which are compatible with high flow and torrential currents. Moroccan rivers would drag vegetation islets (floating rafts) to the Atlantic Ocean where the material would be transported to the nearby coasts of the Canarian archipelago by the trade winds and the marine currents ( Alonso et al. 2000), a process which may be primarily responsible for snail dispersion from Africa to the Canaries.
Thus, Napaeus is almost certainly a genus with northwest African natural affinities while Macaronapaeus most likely displays European natural affinities. Napaeus could be related to Merdigera Held, 1837 , from northwest Africa and southern Europe, or Mauronapaeus Kobelt, 1899 , from the north African countries of Algeria and Tunisia, because both genera are geographically close to Napaeus and all three share a similar type of genital system, including the presence of a penial appendix ( Schileyko 1998, figures 266 and 287, respectively).
Macaronapaeus , which lacks a penial appendix, could have derived from a European population of a penial appendix-bearing, ancestor of genera such as Merdigera or Zebrina Held, 1837 (present in the Iberian Peninsula). Alternatively, Macaronapaeus could also be related to a penial appendix-absent genus present in the Iberian Peninsula, such as Chondrula Beck, 1837 .

| CML |
Universidad Nacional de Tucuman, Coleccion de Mamiferos Lillo (Argentina) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.