
Phyllodoce longipes Kinberg, 1866
|
publication ID |
https://doi.org/10.11646/zootaxa.4924.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:8C98968D-AAF8-403C-AFCC-381B2CC76844 |
|
DOI |
https://doi.org/10.5281/zenodo.4560514 |
|
persistent identifier |
https://treatment.plazi.org/id/3F5087CA-3F32-6700-FF5C-0054FA983C9C |
|
treatment provided by |
Plazi |
|
scientific name |
Phyllodoce longipes Kinberg, 1866 |
| status |
|
Phyllodoce longipes Kinberg, 1866 View in CoL
Figures 20–21 View FIGURE 20 View FIGURE 21
Phyllodoce longipes Kinberg, 1866: 241 View in CoL , figs. 28–30.— Ehlers 1901: 72, Pl. 7, figs. 1–4.—Bergstr̂m 1914: 149, fig. 47.— Day 1963: 394, figs. 3D–F; 1967:144, figs. 5.2A–C.— Pleijel 1990: 146–147, fig. 5 (synonymy). — Blake 2001: 168–169, fig. 4.28.
Anaitides longipes Hartman 1968: 229 View in CoL , figs. 1–3.— Day 1967: 144, fig. 5.2A–C.— Gardiner 1976: 115, fig. 7. — Kravitz & Jones 1979: 15–16. — Gathof 1984: 19–37, figs 19–32.— Parker 1987: 193–194.— Hernández-Alcántara 1992: 156.
Anaitides ( Anaitides) longipes McCammon & Montagner 1979: 357–359 View in CoL , fig. 3. Non Ushakov & Wu 1959.
Phyllodoce papillosa Hyland & Neff 1988 View in CoL : A–3.
Material examined. 27 specimens, length 7.1± 5.2 mm for 45.0± 25.6 segments. Continental shelf in Campos Basin: Hab3 C09 R1 , 23º3’34.2”S 40º41’54.1”W, 1.291 m, 9 May 2008 ( 1 specimen, ZUEC–POL 16325 ); GoogleMaps Hab11 C04 R3 , 22º52’1.9”S 40º57’28.9”W, 92 m, 22 Feb 2009 ( 1 specimen, ZUEC–POL 16364 ); GoogleMaps Hab11 G01 R3 , 21º49’54.7”S 40º44’35.0”W, 29 m, 25 Feb 2009 ( 1 specimen, ZUEC–POL 16663 ); GoogleMaps Hab11 G04 R1 , 22º4’14.4”S 40º6’59.5”W, 91 m, 25 Feb 2009 ( 1 specimen, NHDM–865944); GoogleMaps Hab11 G02 R2 , 21º59’3.7”S 40º25’10.1”W, 52 m, 25 Feb 2009 ( 1 specimen, NHDM–865945); GoogleMaps Hab11 G01 R1 , 21º49’54.4”S 40º44’34.8”W, 25 m, 25 Feb 2009 ( 1 specimen, NHDM–865946); GoogleMaps Hab11 D02 R2 , 22º12’53.0”S 40º51’12.0”W, 52 m, 26 Feb 2009 ( 1 specimen, ZUEC–POL 16365); GoogleMaps Hab11 B02 R1 , 22º37’35.3”S 41º21’51.5”W, 53 m, 27 Feb 2009 ( 1 specimen, ZUEC–POL 16412 ); GoogleMaps Hab11 B01 R3 , 22º37’31.8”S 41º21’51.7”W, 53 m, 27 Feb 2009 ( 1 specimen, NHDM–865947); GoogleMaps Hab11 A02 R1 , 22º56’2.5”S 41º53’50.6”W, 49 m, 28 Feb 2009 ( 1 specimen, ZUEC–POL 16554 ); GoogleMaps Hab13 I01 R2 , 21º11’0.8”S 40º28’27.3”W, 26 m, 5 Mar 2009 ( 1 specimen, ZUEC–POL 16424 ); GoogleMaps Hab13 I04 R3 , 21º9’9.3”S 40º16’5.7”W, 101 m, 7 Mar 2009 ( 1 specimen, ZUEC–POL 16593 ); GoogleMaps Hab13 Foz29 R2 , 21º24’43.5”S 40º25’18.6”W, 33 m, 7 Mar 2009 ( 1 specimen, ZUEC–POL 16537 ); GoogleMaps Hab13 H02 R2 , 21º44’19.3”S 40º17’15.5”W, 49 m, 9 Mar 2009 ( 1 specimen, ZUEC–POL 16400 ); GoogleMaps Hab13 Foz21 R3 , 22º6’21.9”S 40º43’39.4”W, 47 m, 12 Mar 2009 ( 1 specimen, ZUEC–POL 16525 ); GoogleMaps Hab13 Foz41 R2 , 21º45’13.6”S 40º14’7.7”W, 67 m, 14 Mar 2009 ( 1 specimen, ZUEC–POL 16556 ); GoogleMaps Hab13 B01 R1 , 22º41’46.7”S 41º53’46.2”W, 30 m, 16 Mar 2009 ( 2 specimens, ZUEC–POL 16644 ); GoogleMaps Hab13 B01 R2 , 22º41’46.6”S 41º53’46.1”W, 30 m, 16 Mar 2009 ( 1 specimen, ZUEC–POL 16386 ); GoogleMaps Hab13 B01 R3 , 22º41’47.0”S 41º53’46.4”W, 30 m, 16 Mar 2009 ( 1 specimen, ZUEC–POL 16522 ); GoogleMaps Hab16 B04 R3 , 23º10’5.0”S 41º3’7.5”W, 107 m, 2 Jul 2009 ( 1 specimen, ZUEC–POL 16406 ); GoogleMaps Hab16 E04 R2 , 22º17’42.1”S 40º27’0.0”W, 103 m, 4 Jul 2009 ( 1 specimen, ZUEC–POL 16461 ); GoogleMaps Hab16 Foz41 R3 , 21º45’15.3”S 40º14’8.1”W, 66 m, 08 Jul 2009 ( 1 specimen, ZUEC–POL 16395 ); GoogleMaps Hab17 A04 R2 , 23º6’49.6”S 41º55’17.6”W, 110 m, 15 Jul 2009 ( 1 specimen, ZUEC–POL 16437); GoogleMaps Hab 17 Foz 21 R3 , 22º6’20.1”S 40º43’41.0”W, 47 m, 17 Jul 2009 ( 1 specimen, ZUEC–POL 16376 ); GoogleMaps Hab 17 I04 R1 , 21º9’9.7”S 40º16’6.7”W, 103 m, 21 Jul 2009 ( 1 specimen, ZUEC–POL 16662 ); GoogleMaps Hab 17 Foz 02 R1 , 21º21’21.1”S 40º52’9.2”W, 20 m, 23 Jul 2009 ( 1 specimen, ZUEC–POL 16639 ), GoogleMaps Brazil. Additional material examined. Holotype of Phyllodoce longipes , shallow subtidal, Valparaiso, Chile (SNMH–6726) GoogleMaps .
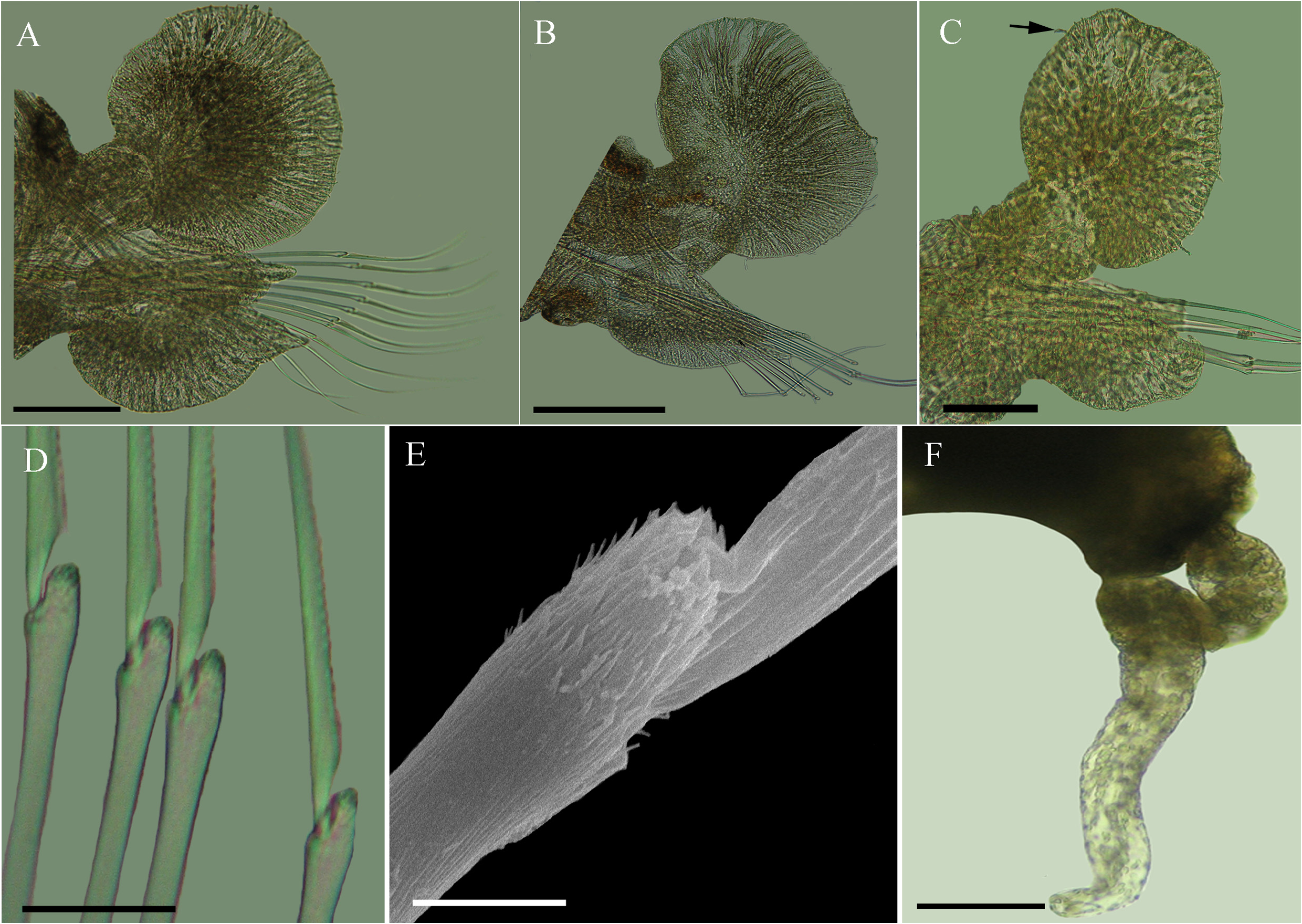
Diagnosis. Proboscis divided into proximal part with 12 longitudinal rows of drop-shaped papillae, six rows on each side, with about 14–15 papillae per row, separated by non-papillated median-dorsal and median-ventral areas and narrow distal area, with large drop-shaped papillae, with acute endings and irregular distribution. Sub-rectangular and median-dorsal cirri with rounded distal ends.
Redescription. Holotype incomplete with 25 segments, 7.0 mm long, 1.0 mm wide median part of body, including parapodia and excluding chaetae. Body long, dorso-ventrally flattened and tapered posteriorly ( Fig. 20A View FIGURE 20 ). Prostomium cordiform, with anterior protuberance and distinct rounded nuchal papilla ( Fig. 20 View FIGURE 20 B–C). Paired frontal, cylindrical antennae and palps with unequal lengths; antennae longer than shorter and robust palps. One pair of epidermal eyes with lenses, reddish and located at median-posterior region of prostomium. Proboscis basally with 12 longitudinal rows of drop-shaped papillae, six rows on each side, with about 14–15 papillae per row, separated by non–papillose median-lateral and median-ventral areas; distal area narrower with large drop-shaped papillae with acute endings and irregular distribution ( Fig. 20A, D View FIGURE 20 ). Terminal ring with 16 papillae alternating between long/ pointed and short/rounded papillae ( Fig. 20 View FIGURE 20 C–B). Segment 1 not visible dorsally. Four pairs of cylindrical tentacular cirri, biarticulated, with short cirrophores and long cirrostyles, located on first three segments ( Fig. 20 View FIGURE 20 A–C). Tentacular cirri of segment 1, reaching segment 4. Dorsal and ventral tentacular cirri of segment 2 reaching segments 9 and 3, respectively. Dorsal tentacular cirri of segment 3 extending to segment 7. Neuropodia and ventral cirri from segment 3. Dorsal cirri with well-developed cirrophores with dorsal extensions, from segment 4; dorsal cirri with distal filaments ( Fig. 21 View FIGURE 21 A–C). Dorsal cirri of anterior segments rounded and symmetrical, those of median and posterior segments asymmetrical, sub-rectangular with rounded distal ends. Anterior and posterior parapodial lobes shorter than dorsal and ventral cirri and median ones longer than ventral cirri, with clear aciculae and bundles of chaetae. Prechaetal lobes bilobate, asymmetrical, with elongated digitiform supracicular lobes, longer than subaciculars. Postchaetal lobes rounded. Ventral cirri horizontally oriented in relation to lobes, from segment 3, asymmetric and dorso-ventrally flattened; rounded anteriorly with tapered distal ends, triangular on median segments and triangular to elongate posteriorly ( Fig. 21 View FIGURE 21 A–C). Compound spinigerous chaetae from segment 4. Rostrum of chaetal shaft surrounded by small denticles; articles with serrated outer edges ( Fig. 21 View FIGURE 21 D–E). Pygidium with one pair of cylindrical, slender anal cirri ( Fig. 21F View FIGURE 21 ).
Colour. Black pigmentation in the anterior part of the prostomium. Distinct black colouration in the first dorsal segments followed by three longitudinal stripes, one median and two lateral ( Fig. 20A View FIGURE 20 ). Black pigmentation also present on parapodial dorsal cirri.
Habitat. Sandy and muddy substrates between 52 and 840 m.
Distribution. Atlantic Ocean, Brazil: Margin of the continental shelf and continental slope in Campos Basin, Rio de Janeiro. Gulf of Mexico, North Carolina, South Africa. Northern Atlantic Ocean in the England Channel. Mediterranean Sea: Denmark, Western Coast of Sweden. Pacific Ocean, Chile ( Pleijel, 1993a).
Remarks. The morphology of the material from the Brazilian coast agrees with the characteristics observed in the examined holotype from Valparaiso, Chile. The affinities between populations from the Atlantic and Pacific coasts of South America can only be confirmed through molecular analyses, which was beyond the scope of this paper. Fauvel (1923) synonymized Phyllodoce macropapillosa Saint-Joseph, 1895 known from Denmark with P. longipes based on similarities in the proboscidial structure and other characteristics, in spite of the different insertion points of chaetae between these species. Anaitis jeffreysii McIntosh, 1908 from Ireland was also synonymized with P. longipes by Parker (1987). In his review of the family, Pleijel (1993) considered very unlikely that the various populations in the United Kingdom and Sweden would be conspecific because of their disjunct geographical distribution. In spite of recognizing this possibility, he identified the specimens collected in Koster, Sweden, as P. cf. longipes . In fact, specimens of P. longipes described for Brazil have dorsal cirri more elongated and a cordiform prostomium that is also anteriorly convex, whereas those referred to as P. cf. longipes from Sweden have median and posterior cirri rounded, although they share the characteristics of a proboscis with a non-papillated dorsal region.
The characteristics considered herein as diagnostic for P. longipes are similar to those specimens described by Ehlers (1901) from Guajatan Bay, Chile and Blake (2001) from North Carolina, USA. Phyllodoce longipes differs from P. rosea , P. colorata sp. nov., P. concava sp. nov., and Phyllodoce sp. B, because these present cuspidate proboscidial papillae ( Blake, 1988; Pleijel, 1993a, b) and from P. bipapillosa sp. nov., Phyllodoce sp. B., P. colorata sp. nov. and P. hiatti by the absence of regular rows of papillae in the proboscis. Phyllodoce longipes has a pair of epidermal eyes whereas P. concava sp. nov., P. lamella sp. nov., P. ovalis sp. nov., and Phyllodoce sp. B lack eyes. Phyllodoce longipes differs the P. tupana sp. nov., Phyllodoce sp. A., Phyllodoce thalia sp. nov., P. brasiliensis sp. nov., P. madeirensis , P. erythrophylla , and P. medipapillata by the presence of a cordiform prostomium anteriorly convex and dorsal cirri with distal filaments.
The morphology of the median sub-rectangular dorsal cirri in P. longipes is similar to P. colorata sp. nov., P. erythrophylla , P. groenlandica , P. cuspidata , P. medipapillata , P. lineata , and P. rosea ; however, it differs from these species by the presence of dorsal cirri terminally rounded on median parapodia. The asymmetry of the lobes in P. longipes is a character shared with P. tupana sp. nov., P. bipapillosa sp. nov., P. thalia sp. nov., P. brasiliensis sp. nov., P. colorata sp. nov., P. madeirensis , P. erythrophylla , P. groenlandica , P. lineata , and P. rosea . However, it differs from these in the shape of the supracicular lobes being digitiform, which is a characteristic of the genus Sige Malmgren, 1865 that apparently has evolved independently within the genus Phyllodoce ( Ushakov, 1972) .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Phyllodoce longipes Kinberg, 1866
| Oliveira, Verônica Maria De, Magalhães, Wagner F. & Lana, Paulo Da Cunha 2021 |
Anaitides longipes
| Hernandez-Alcantara, P. 1992: 156 |
| Parker, M. 1987: 193 |
| Gathof, J. M. 1984: 19 |
| Kravitz, M. J. & Jones, H. R. 1979: 15 |
| Gardiner, S. L. 1976: 115 |
| Hartman, O. 1968: 229 |
| Day, J. H. 1967: 144 |
Phyllodoce longipes
| Blake, J. A. 2001: 168 |
| Pleijel, F. 1990: 146 |
| Day, J. H. 1963: 394 |
| Ehlers, E. 1901: 72 |
| Kinberg, J. G. H. 1866: 241 |