
Hyperolius jackie, Dehling, Maximilian, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.212438 |
|
DOI |
https://doi.org/10.5281/zenodo.5622103 |
|
persistent identifier |
https://treatment.plazi.org/id/3F19A372-2223-FF9D-FF71-FE10FDA5A43B |
|
treatment provided by |
Plazi |
|
scientific name |
Hyperolius jackie |
| status |
sp. nov. |
Hyperolius jackie View in CoL sp. n.
Jackie’s Reed Frog
( Figures 1 View FIGURE 1 & 2 View FIGURE 2 )
Holotype: ZMB 77476 (ex JMD 674), adult male, from a natural pond at Karamba (S 2°28'44.28'', E 29°06'44.50'', 1940 m a.s.l.), Nyungwe National Park, Rwanda, collected on 20 March 2011 at 19:25 h by J. M. Dehling.
Paratopotypes: ZMB 77477–77480 (ex JMD 671– 673, 675), four adult males, collected on 19 & 20 March 2011; ZMB 77481 (ex JMD 726), adult male, collected on 3 April 2011; ZMB 77782 (ex JMD 799), adult male, collected on 18 March 2012; ZMB 77783 (ex JMD 800), adult male, collected on 24 March 2012; otherwise same collection details as holotype.
Diagnosis. The new species is assigned to Hyperolius for showing the following characters considered diagnostic for the genus ( Schiøtz 1999, Channing & Howell 2006): Pupil horizontal, skin smooth, tips of fingers and toes expanded, males with disc-shaped gular flap. Hyperolius jackie is distinguishable from its congeners by the combination of the following characters: (1) size small to medium, SUL of adult males 18.8–23.2 mm, (2) head relatively wide, (3) snout acuminate in dorsal view, rounded in profile, (4) canthus rostralis weakly distinct, (5) tympanum externally discernible, (6) dorsal surfaces smooth with few tiny tubercles, (7) finger and toe webbing not extensive, formulae being I 2.25-(2.25–2.75) II (2+–2.25)-(3–3+) III 2.5-(2+–2.25) IV and I (2–2+)-(2+–2.5) II 1.5- 3 III (2–2-)-(3–3+) IV (3–3-)-(1.75–2-) V, respectively, (8) nuptial pads present in males, (9) pectoral and brachial glands absent in males, (10) hind limbs moderately long, (11) dorsum light green in life with very small brown dots, (12) ventral skin transparent, (13) bones blue, visible through skin, (14) advertisement call consisting of a single pulse group, repeated in series of 2–13 calls, pulse group consisting of 10–18 pulses and lasting 54–107 ms, dominant frequency at 2650– 3150 Hz, (15) 16S rRNA sequence differing by at least 4.5% in the uncorrected pdistance from all currently available sequences of other species of the genus.
Description of holotype ( Figures 1 View FIGURE 1 , 2 View FIGURE 2 ). Measurements are provided in Table 1 View TABLE 1 . Body moderately sturdy, widest at temporal region, tapering to groin; head large (HL/SUL 0.35, HW/SUL 0.36), slightly wider than long (HW/ HL 1.03); snout long (SL/HL 0.48), acuminate in dorsal view, rounded in profile, slightly projecting beyond lower jaw, wider than long (SL/EE 0.80); canthus rostralis weakly distinct between eye and nostril, straight-lined; loreal region oblique, slightly concave; nostrils rounded, directed anterolaterally; situated closer to tip of snout than to eye (EN/NS 1.34), separated from each other by distance subequal to distance between eye and nostril (NN/EN 0.97); eyes directed anterolaterally, protruding, large (ED/HL 0.39), its diameter shorter than snout (ED/SL 0.82); interorbital distance wider than upper eyelid (IO/EW 1.61) and greater than internarial distance (IO/NN 1.25); tympanum concealed under thick layer of skin but discernible, separated from eye by 29% of its diameter; tympanum diameter slightly more than one-third of eye diameter (TD/ED 0.35), upper jaw with dentition; teeth on premaxilla somewhat larger than those on maxilla; choanae small, oval, located far anterolaterally at margins of roof of the mouth, concealed by upper jaw for about the half in ventral view; vomer processes and teeth absent; tongue long (5.6 mm) and broad (4.0 mm at widest point), bilobed for slightly less than one-third of its length, free distally for half its length; median lingual process absent; vocal sac single, median, subgular; gular flap medially arranged as subcircular area of thickened, glandular skin; vocal sac aperture on each side of the mouth, slit-like, long, directed posterolaterally, situated closer to eustachian tube openings than to base of tongue.
Dorsal surfaces of head, trunk and limbs smooth with few scattered tiny low, pointed tubercles; ventral side of limbs and gular region smooth, chin and abdomen slightly more areolate; supratympanic fold absent.
Fore limbs slender; hand moderately large (HND/SUL 0.28); tips of fingers enlarged into broad oval disks, each with circummarginal groove; relative length of fingers: I <II <IV <III; subarticular tubercles rounded, well developed, numbering one on Fingers I and II, two on Fingers III and IV, with proximal tubercles on Fingers III and IV much smaller than distal ones; webbing formula of the hand I 2.25-2.5 II 2 +-3+ III 2.5-2.25 IV; thenar tubercle distinct, about one-sixth the size of base of thumb; palmar tubercles absent; nuptial pad as faintly discernible callosity with few loosely scattered, tiny pointed tubercles on preaxial half of the dorsal surface of Finger I.
Hind limbs slender, moderately long (LEG/SUL 1.65); tibio-tarsal articulation reaching to level of nostrils when legs are adpressed forwardly to body; tibiofibula moderately long (TFL/SUL 0.52), longer than thigh (TFL/THL 1.06); heels overlapping each other slightly with knees flexed and thighs held at right angle to median plane; foot shorter than tibiofibula (FOT/TFL 0.84); relative length of toes: I <II <III <V <IV; discs of toes smaller than those of fingers; subarticular tubercles numbering one on Toes I and II, two on Toes III and V, and three on Toe IV; pedal webbing formula I 2 -2.5 II 1.5- 3 III 2- 3 IV 3-1.75 V; inner metatarsal tubercle small (0.9 mm), 27% length of Toe I (3,3 mm), oval, little prominent; outer one almost circular, low and less distinct, covering larger area than inner one.
Colouration in life. Dorsal side of head, trunk, forelimbs, tibiofibula and tarsus light green with very small brown dots; thigh unpigmented with a yellowish-green dye; ventral side superficially unpigmented, translucent; chin appearing green; gular region and anterior part of chest including lateral sides around insertion of forelimbs, groin, and areas below pelvis, knee, and tibio-tarsal articulation greenish blue; posterior part of chest and abdomen yellow, ventral side of limbs yellowish-green; dorsal side of toes, finger tips and toe tips, and toe webbing greenishyellow; dots on dorsum brown to coppery, loosely scattered on trunk and limbs, most densely around nostril, along canthus rostralis and on upper eyelid; few spots on trunk and tibia slightly enlarged; whitish mottling along edges of both upper and lower jaw, along the border of pigmented dorsal side with unpigmented ventral side, on postaxial sides of forelimb and tarsus and preaxial side of tibiofibula, in infrananal region, and at tibio-tarsal articulation; iris coppery; larger bones (femur, tibiofibula, tarsus, humerus, radioulna, bones of hand and foot) blue, visible through skin ( Figure 2 View FIGURE 2 ).
Colouration in preservative. All colours have faded to white except dark dorsal dots and spots. Larger bones still have a bluish dye.
Variation. Two of the paratypes (ZMB 77477, 77479) had a yellowish-green dorsum at night. Two of the smaller paratypes (ZMB 77477, 77480) have a narrow dorsolateral band running from the posterior edge of the upper eyelid to about halfway between the insertion of arm and the insertion of leg ( Figure 2 View FIGURE 2 ). A dorsolateral band is faintly visible in the smallest (ZMB 77782) and a somewhat larger paratype (ZMB 77478) and lacking in a medium-sized (ZMB 77783) and the two largest paratypes (ZMB 77479, 77481). This might indicate that the band is present in young males and lost with aging. Variation of webbing formula is I 2.25-(2.25–2.75) II (2+–2.25)- (3–3+) III 2.5-(2+–2.25) IV for the hand and I (2–2+)-(2+–2.5) II 1.5- 3 III (2–2-)-(3–3+) IV (3–3-)-(1.75–2-) V for the foot. Measurements of the paratypes are provided in Table 1 View TABLE 1 .
Genetics. Comparison of the mitochondrial 16S ribosomal RNA confirmed that Hyperolius jackie is not conspecific with any of the described taxa for which homologous information is available. Uncorrected p-distance between Hyperolius jackie sp. n. and phylogenetically or geographically close species ranged from 4.5 % to 16.8 % ( Table 2 View TABLE 2 ). The sequences of the two paratypes differed by 0.4 %, reflecting 3 substitutions in the 487 base pairs. A preliminary phylogenetic analysis showed that H. jackie forms a well-supported clade with H. castaneus Ahl, 1931 , H. cystocandicans Richards & Schiøtz, 1977 , H. discodactylus Ahl, 1931 (and H. alticola Ahl, 1931 ), H. frontalis Laurent, 1940 , and H. lateralis Laurent, 1940 (unpublished data). Relationships within this clade are not well resolved but H. jackie appears to be sister to a clade consisting of H. castaneus , H. frontalis , and H. cystocandicans (unpublished data).
Advertisement call. All male type specimens were encountered while calling. Calling started at dusk. The advertisement call of the holotype (ZMB 77476), two paratypes (ZMB 77480 and 77782) and three non-collected specimens were recorded. Air temperature ranged between 15.8 °C and 16.3 °C. The advertisement call consisted of a single pulse group. Usually, a series of calls was emitted, consisting of two to thirteen consecutive calls ( Figure 3 View FIGURE 3 ). The calls were emitted at a rate of 5.54 ± 0.35 (4.81–6.29) per second (mean ± SD [range]) within a series. The pulse group was composed of 10–18 (median: 14) pulses and had a length of 75.4 ± 12.8 (54–107) ms ( Figure 3 View FIGURE 3 ). Pulse rate varied between notes and also within the note between 125 and 250 pulses per second, being highest at the beginning of the note and lowered towards the end. Amplitude modulation was prominent. Relative amplitude rose from the beginning to the middle of the call and declined towards the end. Dominant frequency ranged between 2650 and 3150 Hz ( Figure 3 View FIGURE 3 ). There were prominent harmonics at ca. 6000, 9000, 12000 and 15000 Hz, and less distinct also at 18000 and 21000 Hz. There was a slight frequency modulation with the dominant frequency of the single pulses within a note differing for up to 100 Hz but without an obvious pattern. In most calls, however, the lowest frequencies were measured in the first half of the pulses and the highest in the second half. About two-thirds of the call series were initiated by another call type which preceded the advertisement calls. It consisted of a single pulse group and generally resembled the advertisement call in structure and dominant frequency but was much longer (180.1 ± 28.7 [130–223] ms), consisted of a higher number of pulses (17–25; median 20), and had a much lower pulse repetition rate (90–166 pulses per second) ( Figure 3 View FIGURE 3 ). This call appeared to be an abbreviated aggression call. Three aggression calls, which were emitted without adding a series of advertisement calls, were recorded and analysed. They were longer (244–341 ms), consisted of 26–35 pulses, and had a pulse repetition rate of 71–142 pulses per second.
Ecology and distribution. Type specimens were collected between 19:00 h and 00:45 h perching on leaves in vegetation ( Rubus steudneri and Juncus effusus ), about 1 m above the ground. Air temperatures recorded during collection with males being active ranged from 15.0°C (19 March 2011, 22:30 h) to 18.2°C (20 March 2011, 19:00h). All type specimens were collected at a single locality, a swamp at Karamba in the southwestern part of the Nyungwe National Park ( Figure 4 View FIGURE 4 ). I heard and recorded the characteristic call of the species at another locality at a small stream between the type locality and the edge of the forest (S 2°27'49.81'', E 29°06'02.88'', 1881 m a.s.l.) but did not collected nor see specimens there ( Figure 4 View FIGURE 4 ). The species probably also occurs in similar habitats in other areas of Nyungwe National Park and in the adjacent Kibira National Park in Burundi. I found four frog species syntopically with the new species, Hyperolius castaneus , Leptopelis kivuensis Ahl, 1929 , Amietia cf. angolensis (Bocage, 1866) , and Xenopus wittei Tinsley, Kobel and Fischberg, 1979 . Phlyctimantis verrucosus Boulenger, 1912 has also been recorded at this locality (E. Fischer, pers. comm.). At the second site at the stream, several calling males of Hyperolius discodactylus were recorded. Apparently, Hyperolius jackie is quite sensitive to light. Shortly after I started to look for specimens around the pond at Karamba with a torch, males stopped calling and remained silent even after I turned off the light. In contrast, specimens of Hyperolius castaneus kept calling even if the light beam was pointed directly at them from close distance.
Etymology. The species name is dedicated on behalf of Gordon Buchan to Jacqueline Mary Buchan in recognition of support of biodiversity research and nature conservation through the BIOPAT initiative. The name is used as an invariable noun in apposition.


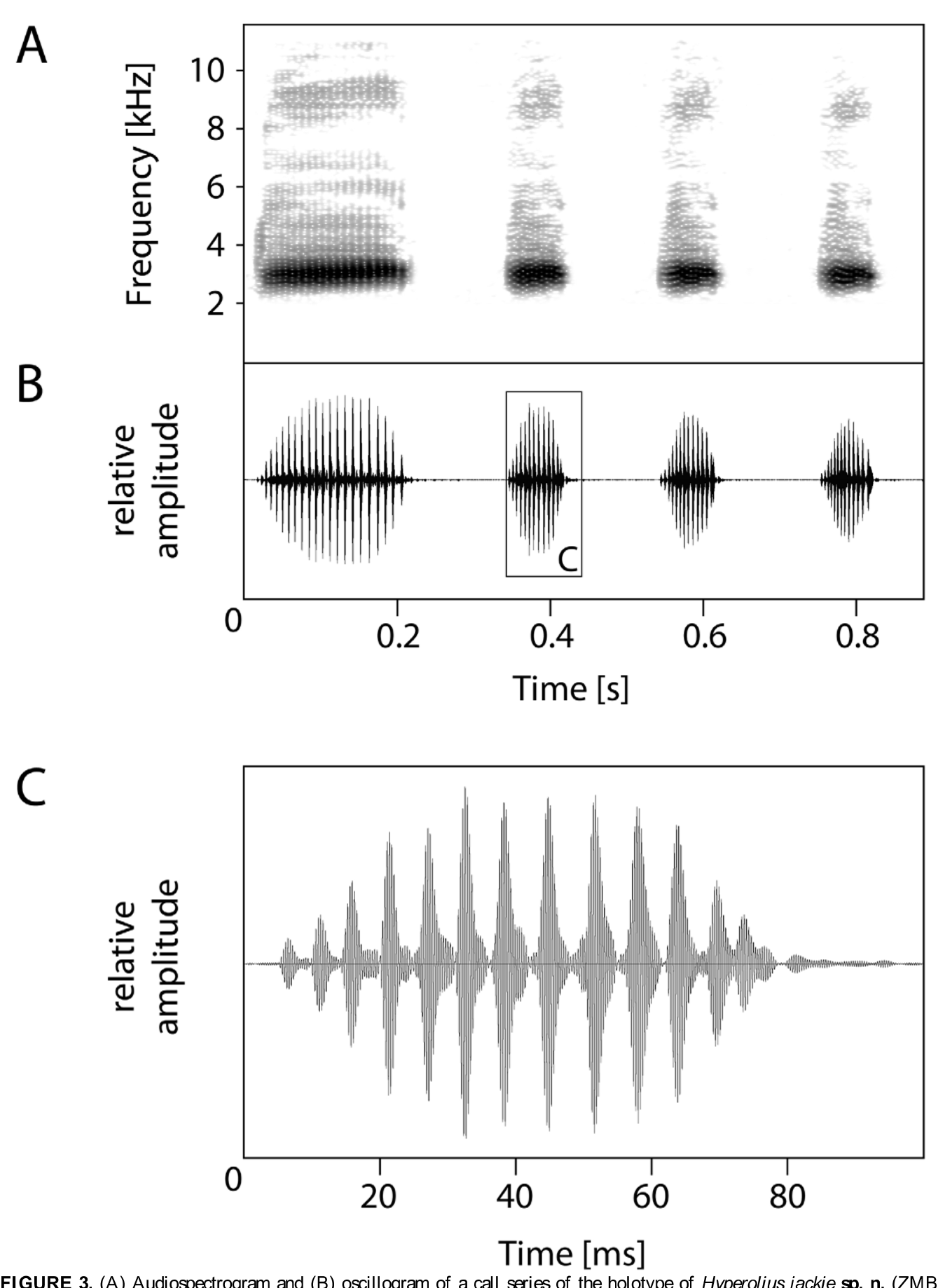
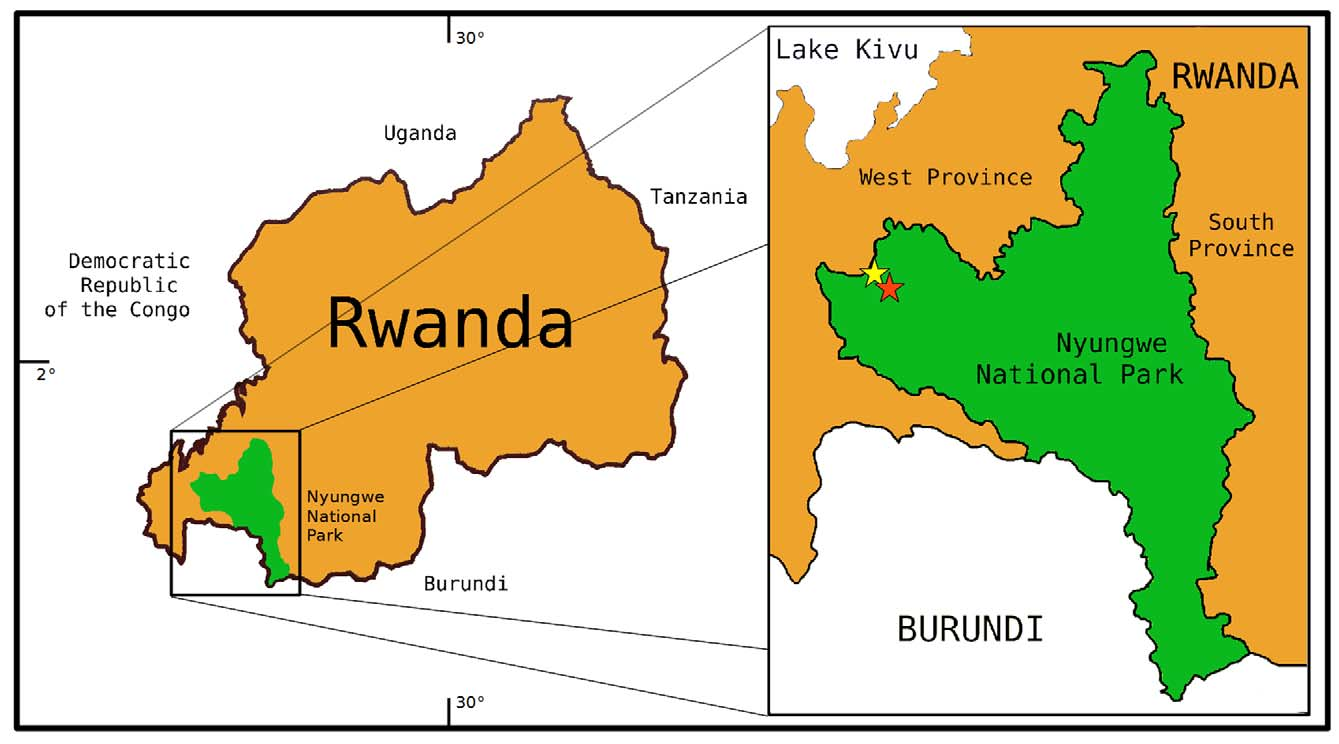
TABLE 1. Measurements and proportions of the type series of Hyperolius jackie sp. n. For abbreviations see Material & Methods. ZMB 77476 77477 77478 77479 77480 77481 77782 77783 Status Holotype Paratype Paratype Paratype Paratype Paratype Paratype Paratype
| SUL 23.1 | 20.1 | 21.2 | 23.2 | 19.0 | 21.5 | 18.8 | 19.8 |
|---|---|---|---|---|---|---|---|
| TFL 12.1 | 10.2 | 10.8 | 11.8 | 9.7 | 10.8 | 9.1 | 9.9 |
| LEG 38.2 | 32.3 | 33.7 | 37.6 | 30.8 | 34.9 | 28.8 | 32.0 |
| TarL 16.6 | 14.1 | 14.8 | 16.9 | 14.0 | 15.8 | 13.3 | 14.1 |
| THL 11.4 | 10.0 | 10.1 | 11.0 | 9.3 | 10.5 | 8.4 | 9.6 |
| FOT 10.1 | 8.4 | 8.8 | 10.3 | 8.3 | 9.9 | 8.1 | 8.9 |
| HND 6.5 | 5.8 | 6.0 | 6.9 | 5.3 | 6.8 | 5.7 | 5.7 |
| HW 8.3 | 7.5 | 7.5 | 8.2 | 7.0 | 8.1 | 7.0 | 7.4 |
| HL 8.1 | 7.2 | 6.9 | 8.1 | 6.5 | 7.4 | 6.8 | 6.9 |
| ED 3.2 | 2.9 | 2.9 | 3.1 | 2.6 | 2.9 | 2.8 | 2.8 |
| TD 1.1 | 1.1 | 1.1 | 1.1 | 0.9 | 1.0 | 0.9 | 1.0 |
| ET 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 |
| EN 2.3 | 1.9 | 1.9 | 2.3 | 1.8 | 2.1 | 1.7 | 1.8 |
| NS 1.7 | 1.6 | 1.6 | 1.7 | 1.5 | 1.8 | 1.3 | 1.5 |
| NN 2.3 | 2.1 | 2.2 | 2.3 | 2.0 | 2.3 | 1.9 | 2.0 |
| SL 3.8 | 3.1 | 3.2 | 3.8 | 3.0 | 3.6 | 2.9 | 3.0 |
| EE 4.8 | 4.2 | 4.2 | 5.0 | 4.1 | 4.7 | 4.1 | 4.1 |
| IO 2.8 | 2.4 | 2.5 | 2.7 | 2.3 | 2.6 | 2.3 | 2.5 |
| EW 1.7 | 1.7 | 1.6 | 1.8 | 1.7 | 1.7 | 1.7 | 1.6 |
| GF 6.0 | 5.7 | 5.8 | 6.0 | 5.2 | 5.3 | 5.5 | 5.4 |
| TFL/SUL 0.52 | 0.51 | 0.51 | 0.51 | 0.51 | 0.50 | 0.48 | 0.50 |
| FOT/SUL 0.44 | 0.42 | 0.42 | 0.44 | 0.44 | 0.46 | 0.43 | 0.45 |
| LEG/SUL 1.65 | 1.61 | 1.59 | 1.62 | 1.62 | 1.62 | 1.53 | 1.61 |
| THL/TFL 0.94 | 0.98 | 0.94 | 0.93 | 0.96 | 0.97 | 0.93 | 0.97 |
| FOT/TFL 0.84 | 0.82 | 0.82 | 0.88 | 0.86 | 0.91 | 0.89 | 0.90 |
| HND/SUL 0.28 | 0.29 | 0.28 | 0.30 | 0.28 | 0.31 | 0.30 | 0.29 |
| HW/SUL 0.36 | 0.37 | 0.36 | 0.36 | 0.37 | 0.37 | 0.37 | 0.37 |
| HL/SUL 0.35 | 0.36 | 0.33 | 0.35 | 0.34 | 0.34 | 0.36 | 0.35 |
| HW/HL 1.03 | 1.03 | 1.09 | 1.02 | 1.06 | 1.09 | 1.04 | 1.08 |
| SL/HL 0.48 | 0.43 | 0.46 | 0.47 | 0.45 | 0.48 | 0.42 | 0.43 |
| SL/EE 0.80 | 0.73 | 0.77 | 0.77 | 0.73 | 0.75 | 0.70 | 0.72 |
| IO/NN 1.25 | 1.12 | 1.18 | 1.17 | 1.17 | 1.16 | 1.17 | 1.25 |
| IO/EW 1.61 | 1.40 | 1.55 | 1.46 | 1.37 | 1.53 | 1.31 | 1.56 |
| TD/ED 0.35 | 0.38 | 0.37 | 0.37 | 0.34 | 0.34 | 0.32 | 0.35 |
| EN/NS 1.34 | 1.16 | 1.22 | 1.34 | 1.22 | 1.20 | 1.28 | 1.18 |
| NN/EN 0.97 | 1.10 | 1.14 | 1.00 | 1.12 | 1.07 | 1.15 | 1.10 |
| ED/SL 0.82 | 0.95 | 0.91 | 0.80 | 0.89 | 0.82 | 0.97 | 0.93 |
| ED/HL 0.39 | 0.40 | 0.42 | 0.38 | 0.40 | 0.39 | 0.41 | 0.40 |
| ED/EN 1.36 | 1.53 | 1.54 | 1.32 | 1.49 | 1.37 | 1.64 | 1.52 |
| IO/HW 0.34 | 0.32 | 0.34 | 0.33 | 0.34 | 0.33 | 0.32 | 0.34 |
TABLE 2. Uncorrected p-distance between Hyperolius jackie sp. n. (paratype ZMB 77481, GenBank accession number JQ 966571) and other species of Hyperolius from Rwanda and adjacent areas, based on 487 base pairs of mitochondrial 16 S ribosomal RNA.
| Species | Origin | GenBank # | Voucher p-distance [%] |
|---|---|---|---|
| Hyperolius jackie (paratype) | Nyungwe National Park, Rwanda | ------------ | ZMB 77480 0.4 |
| Hyperolius castaneus | Nyungwe National Park, Rwanda | JQ423936 View Materials | ZMB 77537 4.5 |
| Hyperolius cf. cinnamomeoventris | Butare, Rwanda | JQ966568 View Materials | ZMB 77533 9.6 |
| Hyperolius cystocandicans | Mt. Kenya, Kenya | FJ594079 View Materials | ZFMK 77611 6.6 |
| Hyperolius discodactylus | Nyungwe National Park, Rwanda | JQ966565 View Materials | ZMB 77536 6.4 |
| Hyperolius alticola (considered a synonym of H. discodactylus ) | Bwindi Impenetrable National Park, Uganda | DQ283225 View Materials | CAS 202047 7.1 |
| Hyperolius frontalis | data not provided | AY603986 View Materials | CAS 201986 5.8 |
| Hyperolius kivuensis | Butare, Rwanda | JQ966567 View Materials | ZMB 77532 10.3 |
| Hyperolius lateralis | Butare, Rwanda | JQ966569 View Materials | ZMB 77534 7.5 |
| Hyperolius sp. (viridiflavus- group) | Gitarama, Rwanda | JQ966570 View Materials | ZMB 77535 14.5 |
| Hyperolius sp. (viridiflavus- group) | Mizingo, Rwanda | JQ966566 View Materials | ZMB 77531 14.8 |
| Hyperolius sp. ( nasutus -group) | Butare, Rwanda | ------------ | ZMB 77141 16.8 |
| ZMB |
Museum für Naturkunde Berlin (Zoological Collections) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |