
Goezeella siluri Fuhrmann, 1916
|
publication ID |
https://doi.org/10.5281/zenodo.893547 |
|
DOI |
https://doi.org/10.5281/zenodo.5672077 |
|
persistent identifier |
https://treatment.plazi.org/id/3B7D7060-FFB0-E77C-FC27-2D272EEAF9DA |
|
treatment provided by |
Plazi |
|
scientific name |
Goezeella siluri Fuhrmann, 1916 |
| status |
|
Goezeella siluri Fuhrmann, 1916
Figs 3, 4, 10-19 View Figs 1 - 10 View Figs 11 - 14 View Fig. 15 View Figs 16 - 19
Goezeella siluri Fuhrmann, 1916: 385 .
Monticellia siluri . – Woodland, 1925: 714.
Corallobothrium siluri . – Harwood, 1933: 140.
Goezeella piramutab Woodland, 1933a: 488 .
Monticellia piramutab . – Woodland, 1935: 222.
Spatulifer piramutab . – Brooks & Deardorff, 1980: 17.
Spatulifer siluri . – Brooks, 1995: 365.
Material examined
Syntype: MHNG-PLAT-36375, a whole-mounted specimen (2 slides) and 12 slides of serial cross-sections. Additional specimens: MHNG-PLAT-19858, a wholemounted specimen (4 slides) and 6 slides of serial crosssections, collected on 13.04.1992, host field no. Br 237a. – CHIOC 38858, a whole-mounted specimen (one slide) collected on 30.09.1995, host field no. Br 436. – MHNG- PLAT-21845, 21877, 2 uncomplete whole-mounted specimens (3 slides) and 3 slides of serial cross-sections collected on 0 2.09.1995, host field no. Br 472. – MHNG- PLAT-21879, CHIOC 38859, a whole-mounted specimen (4 slides) and 6 slides of serial cross-sections collected on 19.10.1995, host field No. Br 811; all specimens from P. pirinampu from Itacoatiara, State of Amazonas, Brazil (3°09’S, 58°26’W), collected by A. A. Rego and A. de Chambrier. – MHNG-PLAT-85161, a whole-mounted specimen (3 slides), collected on 0 7.10.2011, host field no. PI 819a; hologenophore; specimen from P. pirinampu from Iquitos, Region of Loreto, Peru (3°47’S, 73°20’W).
Type locality: Amazon River basin (specific locality unknown).
Other localities: Delta of the Orinoco River, Venezuela; Amazon River, Itacoatiara, State of Amazonas, Brazil; Amazon River, Iquitos, Region of Loreto, Peru.
Type host: Blue whale catfish Cetopsis coecutiens (Lichtenstein, 1819) ( Siluriformes : Cetopsidae ).
Other definitive hosts: Cetopsis othonops (Eigenmann) ( Siluriformes : Cetopsidae ) and Brachyplatystoma vaillantii (Valenciennes) ( Siluriformes : Pimelodidae ) (doubtful host – see remarks below); Pinirampus pirinampu (Spix & Agassiz) ( Siluriformes : Pimelodidae ).
Site of infection: Anterior intestine.
Prevalence of infection: Fuhrmann (1916) found 4 tapeworms in one C. coecutiens , but did not provide the total number of fish examined; 43 P. pirinampu examined/8 fish infected (19%) from Itacoatiara, Brazil.
Morphological description: Fuhrmann Chambrier et al. (2004a), present study.
(1916),
de Representative DNA sequences: A fragment of 1491 bp long of the lsr DNA (D1–D3 domains) (GenBank MF370207 View Materials ). The newly generated sequence of an isolate of G. siluri from P. pirinampu is 483 bp longer than the sequence from the same host and river basin available in the GenBank ( AJ388612 View Materials ).
Redescription (based on 6 whole worms, serial crosssections of mature proglottids and 1 scolex studied using SEM from P. pirinampu ; measurements taken from the syntype in brackets): Proteocephalidae . Testes, ovary, vitelline follicles and uterus cortical; medium-sized worms. Total body length 90-230 mm (n = 3), maximum width up to 3 mm (n = 3). Strobila acraspedote, anapolytic, with longitudinal and transverse grooves, consisting of about 335-360 proglottids: 195-200 immature (up to appearance of spermatozoa in vas deferens), 45-55 mature (up to appearance of eggs in uterus), 50-55 pregravid (up to appearance of hooks in oncospheres) and 45-50 gravid. Immature and mature proglottids much wider than long (length: width ratio 0.10-0.20), pregravid proglottids wider than long (length: width ratio 0.45-0.55), gravid proglottids wider than long to longer than wide (length: width ratio 0.80-1.93).
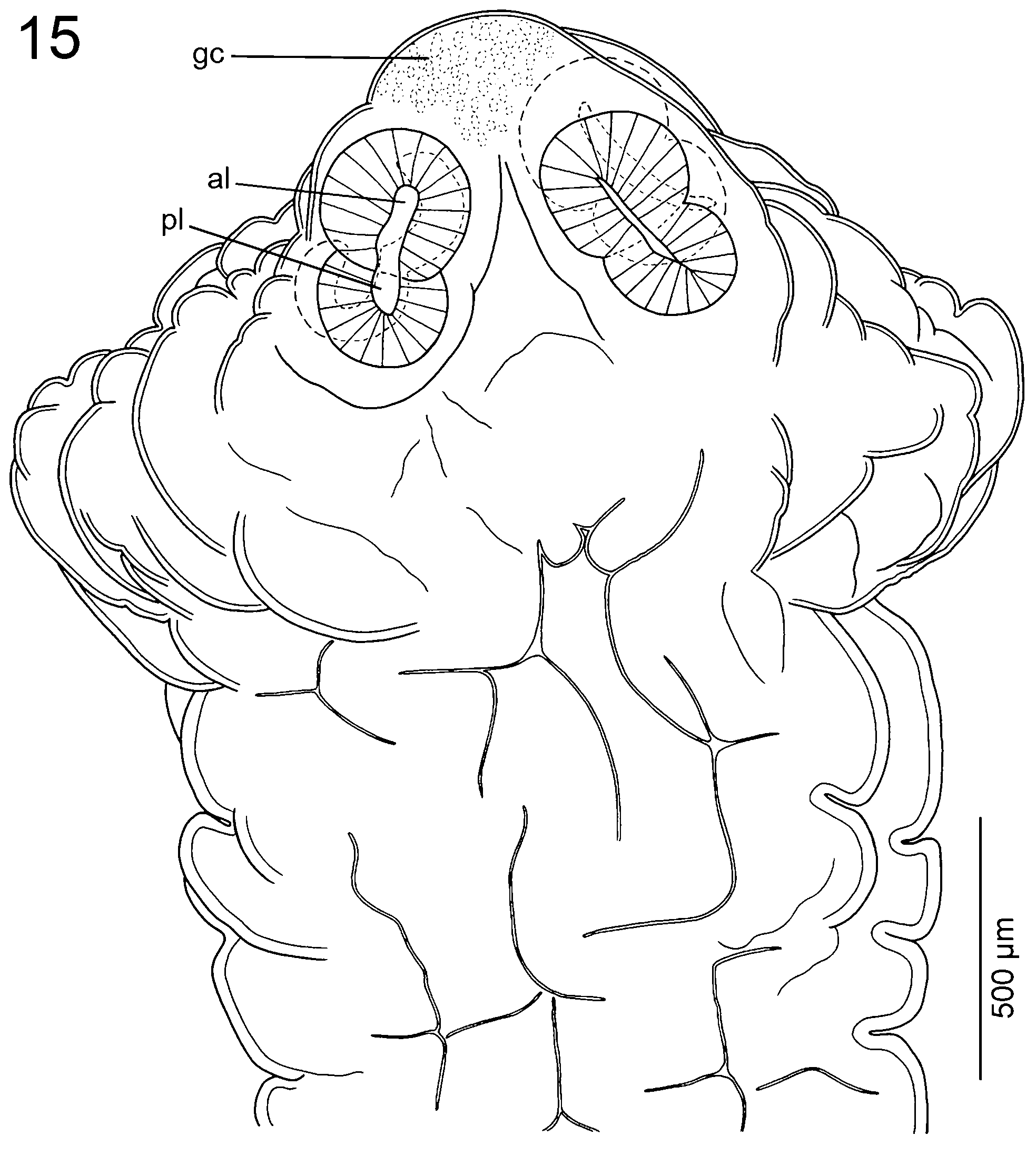
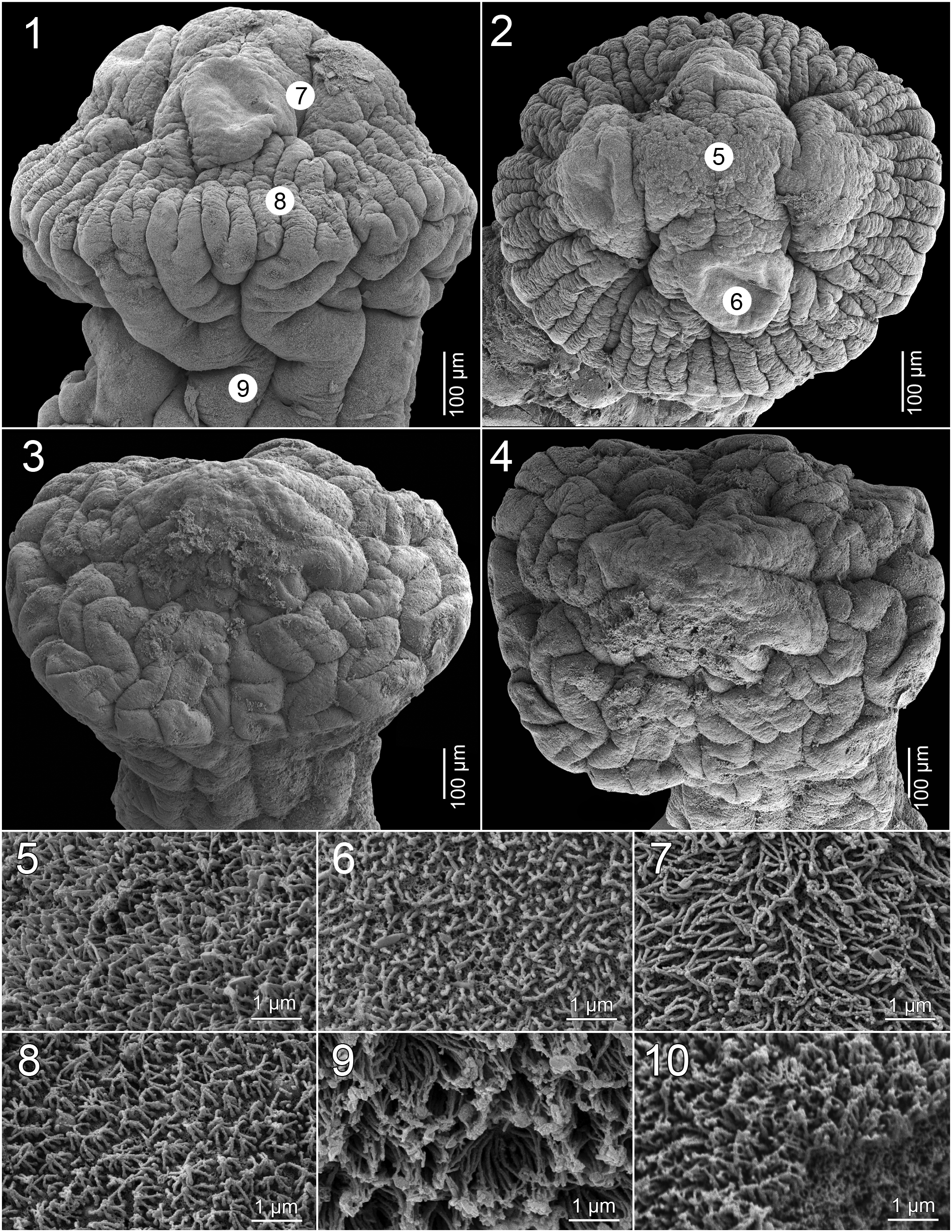
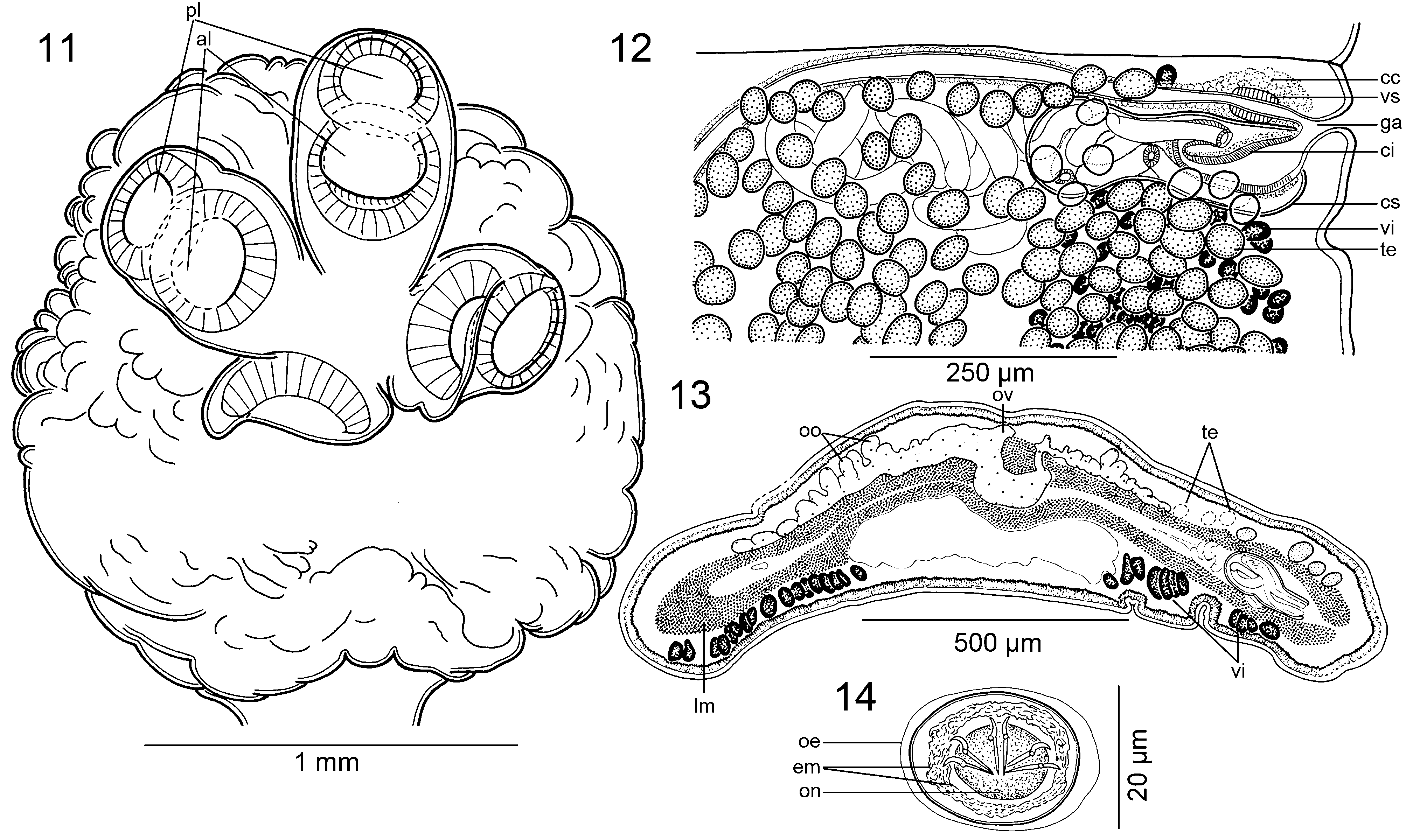
Scolex 1.18-1.60 × 1.45-1.94 mm (n = 5), much wider than neck (proliferation zone), 1.77-1.80 × 1.40- 1.45 mm, bearing 4 robust suckers, biloculate, with loculi variable in size; anterior loculus 210-295 (x = 255; n = 5) in diameter, posterior loculus 205-255 (x = 217; n = 5) in diameter; conspicuous septum separating each loculus ( Figs 11 View Figs 11 - 14 , 15 View Fig. 15 ). Metascolex present, uniformly wrinkled. Apex conical, lacking apical organ, with numerous gland-cells ( Figs 3, 4 View Figs 1 - 10 , 11 View Figs 11 - 14 , 15 View Fig. 15 ). Apex of scolex and lumen of suckers covered with acicular filitriches, similar in density (not shown); surface between suckers, base of metascolex and neck covered with capilliform filitriches, similar in density (not shown); pregravid proglottids covered with acicular filitriches ( Fig. 10 View Figs 1 - 10 ).
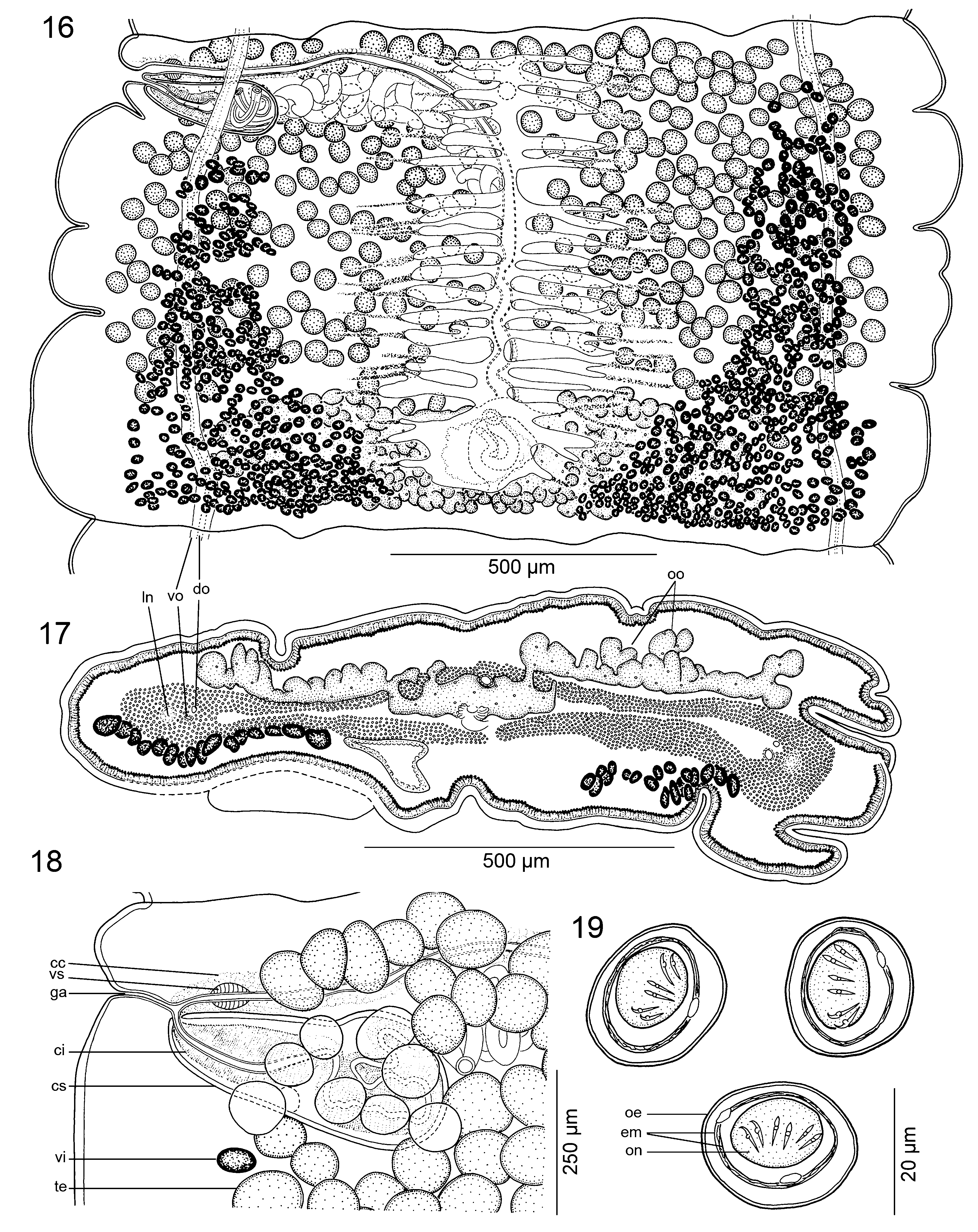
Inner longitudinal musculature well-developed, composed by numerous, individual muscle fibres, concentrated on lateral sides of proglottids ( Figs 13 View Figs 11 - 14 , 17 View Figs 16 - 19 ). Osmoregulatory canals situated at same level, median to lateral-most testes and vitelline follicles, almost straight ( Figs 16, 17 View Figs 16 - 19 ); ventral osmoregulatory canal wider than dorsal one ( Figs 16, 17 View Figs 16 - 19 ).
Testes numerous, spherical, small, 55-80 in diameter, in 1 or 2 irregular layers, 282-366 (x = 314; n = 13) [380- 430] per mature proglottids ( Figs 12 View Figs 11 - 14 , 16, 18 View Figs 16 - 19 ). Testes form 1 irregular field on dorsal side, less numerous in median line of proglottids (uterine stem), usually surpassing osmoregulatory canals, dorsally overlapping cirrus-sac, vitelline follicles and sometimes ovary ( Figs 12, 13 View Figs 11 - 14 , 16, 18 View Figs 16 - 19 ). Testes present also in gravid proglottids.
Vas deferens coiled, with loops forming elongated field reaching to, but not crossing, median line of proglottid ( Figs 12 View Figs 11 - 14 , 16, 18 View Figs 16 - 19 ). Cirrus-sac pear-shaped, thin-walled, slightly widened towards distal (terminal) part ( Figs 12, 13 View Figs 11 - 14 , 16, 18 View Figs 16 - 19 ), 220-340 × 95-145 (n = 13) [250-300 × 80- 110], its length representing 10-21% (x = 15; n = 13) [14-25%] of proglottid width. Sperm duct (internal vas deferens) sinuous. Cirrus muscular, reaching up to 82% (n = 13) [50%] of cirrus-sac length. Common genital atrium narrow, deep ( Figs 12, 13 View Figs 11 - 14 , 16, 18 View Figs 16 - 19 ). Genital pores alternating irregularly, markedly pre-equatorial, situated at 3-10% (x = 6; n = 13) [5-14%] of proglottid length from anterior margin ( Figs 12 View Figs 11 - 14 , 16, 18 View Figs 16 - 19 ).
Ovarywithwideisthmusinmedullaand 2 follicular, grapelike lobes penetrating inner longitudinal musculature to dorsal cortex; numerous dorsal outgrowths present ( Figs 13 View Figs 11 - 14 , 17 View Figs 16 - 19 ). Length of ovary represents 27-37% (x = 30%; n = 13) [33-45%] of proglottid length, its width representing 57-76% (x = 68%; n = 13) [74-86%] of proglottid width ( Figs 16, 17 View Figs 16 - 19 ). Mehlis’ gland about 110- 165 in diameter, representing 7-20% of proglottid width (n = 13). Relative ovarian size, i.e., percentage of ovary surface to total surface of mature or pregravid proglottids (see de Chambrier et al., 2012), 11-16% (x = 13%; n = 10).
Vaginal canal slightly sinuous, surrounded by chromophilic cells, wider in terminal part ( pars copulatrix vaginae), possessing a terminal vaginal sphincter ( Figs 12 View Figs 11 - 14 , 16, 17 View Figs 16 - 19 ). Vagina anterior to cirrus-sac (n = 35). Vitelline follicles cortical, ventral, forming 2 long uninterrupted bands, occupying large triangular field, widened and confluent posteriorly at ovary level ( Figs 13 View Figs 11 - 14 , 16, 17 View Figs 16 - 19 ). Length of bands represents 77-94% (x = 86%) [94-98%] and 84-98% (x = 90%; n = 13) [91- 98%] of length of proglottid on poral and aporal side, respectively ( Figs 12 View Figs 11 - 14 , 16 View Figs 16 - 19 ).
Uterus cortical, with development of type 2 (see de Chambrier et al., 2004b, 2015b); uterine stem and diverticula (lateral uterine branches) in mature and pregravid proglottids lined with numerous chromophilic cells, extended much beyond branches ( Fig. 16 View Figs 16 - 19 ). Uterus with 20-26 [17-22] lateral diverticula on each side ( Fig. 16 View Figs 16 - 19 ). Eggs oval, outer envelope 22-23 × 19-20, bilayered embryophore 17-18 × 12-14 [21-22 ×14-22], oncosphere 11-12 × 9-10 [10-11 × 7-8], embryonic hooks 6-7 long ( Fig. 19 View Figs 16 - 19 ).
Remarks: The original description of Goezeella siluri was detailed and contained basic measurements and illustrations ( Fuhrmann, 1916). However, the only complete specimen preserved is slightly decomposed, contracted and twisted on the slide. Moreover, Fuhrmann (1916) overlooked an important feature of the scolex morphology, i.e. the presence of biloculate, rather than uniloculate, suckers (compare the same scolex drawn in his Fig. 2 View Figs 1 - 10 and Fig. 11 View Figs 11 - 14 of the present study). This characteristic was first reported by Rego (1975; see his fig. 8), but not by Woodland (1933a) as reported by Brooks & Deardorff (1980). In fact, morphology of the suckers may be difficult to observe, especially if the suckers are hidden within wrinkles of a contracted metascolex (see Fig. 3 View Figs 1 - 10 ).
The specimens from P. pirinampu are considered conspecific with G. siluri even though they differ from those found in C. coecutiens described by Fuhrmann (1916) in the lower number of the testes (282-366 vs. 380- 430), a longer cirrus (reaching up to 82% of the cirrus-sac length vs. reaching only up to 50%), and in the anterior extent of the bands of vitelline follicles on the poral side, which represents 77-94% of proglottids length vs. 94- 98% in the syntype. These differences are considered to be accounted for by intraspecific variability, but poor quality of the type material of G. siluri should also be considered. Therefore, new material of G. siluri from its type host, C. coecutiens , is needed for confirmation of measurements taken from the type specimen.
Specimens from Pseudocetopsis othonops (syn. of Cetopsis othonops ) in the Orinoco River, Venezuela, reported by Brooks & Rasmussen (1984) as G. siluri , possess the vagina anterior or posterior to the cirrus-sac, unlike the exclusively anterior position in all species of Goezeella including G. siluri and the newly described species (see below). This feature has been broadly used to differentiate species of proteocephalid cestodes (Arredondo & Gil de Pertierra, 2010; Gil de Pertierra & de Chambrier, 2013), but Alves et al. (2017b) described conspecific cestodes (genetically identical individuals based on the lsr DNA) with both the anterior and posterior vagina in relation to the cirrus-sac. Therefore, this character should be considered with caution. Since all specimens from P. othonops were fixed in AFA (alcoholformalin-acetic acid), they could not be used for DNA sequencing.
Brooks & Rasmussen (1984) also reported immature cestodes identified as G. siluri from B. vaillantii , but reliable identification of juvenile specimens is almost never possible because they lack key morphological traits that are present only in their mature forms. Two of the present authors (TS and AdC) have not found G. siluri in any of the 25 B. vaillantii examined (de Chambrier et al., 2015a); therefore, this record of G. siluri in a species of Brachyplatystoma Bleeker requires verification.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Goezeella siluri Fuhrmann, 1916
| Philippe V. Alves, Alain de Chambrier, José L. Luque & Tomáš Scholz 2017 |
Goezeella piramutab
| Woodland 1933: 488 |
Goezeella siluri
| Fuhrmann 1916: 385 |