
Paraloricaria commersonoides ( Devincenzi 1943 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5406.2.11 |
|
DOI |
https://doi.org/10.5281/zenodo.10620972 |
|
persistent identifier |
https://treatment.plazi.org/id/3A668786-FFD6-F761-FF21-FE22A300FE35 |
|
treatment provided by |
Plazi |
|
scientific name |
Paraloricaria commersonoides ( Devincenzi 1943 ) |
| status |
|
Paraloricaria commersonoides ( Devincenzi 1943) View in CoL ( Fig. 1 View FIGURE 1 )
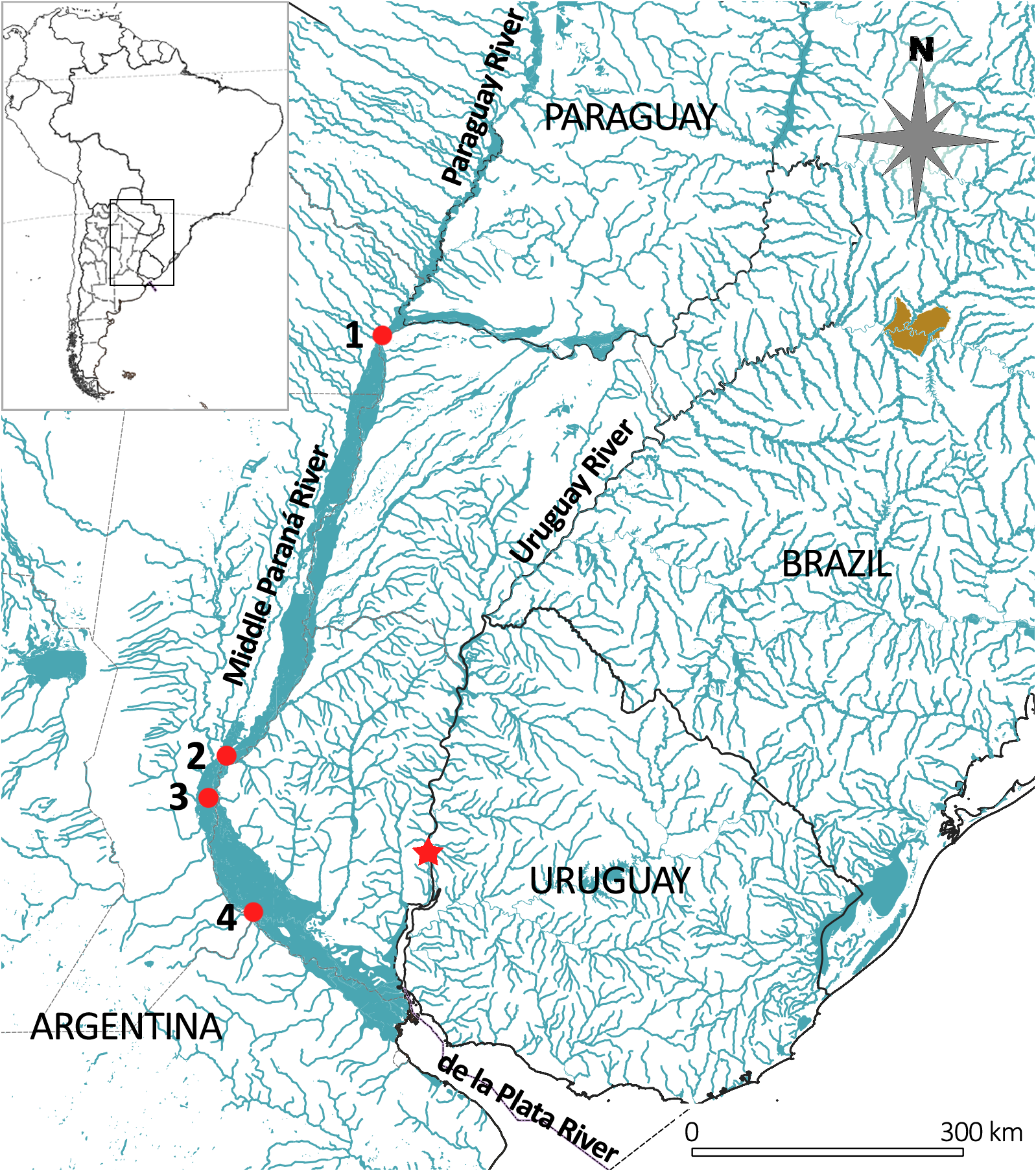
ARGENTINA, BUENOS AIRES Province. MPS-ZI 1546. 1 ex, 230 mm SL. Paraná River near San Nicolás, 33°20’S 060°15’W, col.: A. Scasso. 1950?. GoogleMaps MPS-ZI 1547. 1 ex, 355 mm SL. Paraná River near San Nicolás, 33°15’36,7’’S 060°14’36’’W, col.: G. and T. Carbonell. 03.Jun.2023. GoogleMaps CHACO Province. MPS-ZI 1544. 1 ex, 145.5 mm SL. Paraná River, 27°27’51,1”S 058°52’02,5”W, col.: P. Arrieta, L. Behr, L. Balboni, A. Delgado, D. Fuchs, N. Got, N. Lischetti, G. Picotti, J. Salva, D. Somoza, F. Vargas. 18.Feb.2022. GoogleMaps SANTA FE Province. MCP 14066. 1 ex. 228.5 mm SL. Río Paraná in Santa Fe, 31°35’S 060°41’W (approx.). Col: R. E. Reis. 1990 GoogleMaps .
Morphometric measurements and meristic counts of the analyzed specimens are presented in Table 1 View TABLE 1 . The collected specimens have characteristics that allow their identification as Paraloricaria commersonoides , by comparison with the description of the type material of the species ( Devincenzi 1943).
It can be distinguished ( Table 2 View TABLE 2 ) from the other two species of the genus P. agastor Isbrucker 1979 and P. vetula (Valenciennes 1836) by the dorsal coloration of head and body, light brown with small rounded or elongated dark spots (vs. 4–5 dark broad transverse stripes); by the head longer than wide (head width 70.3–79.8 % vs. 96 % HL), the lower head height (31.4–38.0 % HL vs. 43.0–60.4 % in P. agastor and 38.2–54.6 % HL in P. vetula ) and by the shape of the caudal peduncle, which remains wide until reaching the first lateral plates with fused keels, while in the other two species becomes narrower progressively. This last feature is not as evident in smaller specimens. Measures of P. agastor and P. vetula in Table 2 View TABLE 2 were taken from Rodríguez (2003).
The discovery of new specimens has allowed to improve the knowledge of the morphology of the species. There is a particular conformation of trifurcated premaxillary barbels, located on both sides of the mouth and directed towards the center of it. This character was mentioned by Devincenzi (1943: 4) in the original description, noting that “The tentacles of the upper lip are bi and even trifurcated”, and it is absent in the other two species of the genus. Unlike what is described for the type specimen, it is emphasized that both external rays (dorsal and ventral) of the caudal fin can be prolonged in long filaments with alternating light and dark rings. The other species of the genus also have the upper caudal rays prolonged in a long filament ( Rodríguez 2003). The abdominal plates of the specimens analyzed cover the abdomen almost completely, and they are, in part, small plates that are not in contact with each other. Covain & Fisch-Muller (2007) used this character, which is not clearly mentioned by Devincenzi (1943) or by Isbrücker (1979) in the description of the genus, to differentiate Paraloricaria from the other species of the Loricaria group. However, Rodríguez (2003: 73, 76) mentioned that in both P. agastor and P. vetula these plates “may appear covering the entire abdomen (up to the gill openings), may be missing in some areas or missing, practically, throughout the abdomen.”. In particular, the number of teeth has been somewhat variable. Isbrücker (1979) comments that, due to the phenomenon of tooth replacement, specimens of Paraloricaria can present, at any given time, five teeth or even less in each upper half of the jaw. The typical small dark spots may disappear in long-preserved specimens: MCP 14066 shows no visible spots; on MPS-ZI 1544, preserved almost two years ago, the spots are blurred.
Preliminary analysis of stomach contents shows a benthic diet, with a dominant presence of Diptera larvae of an indeterminate family, close to Chironomidae (Miguel Saigo pers. com.), along with remains of other insects. The congeneric species P. vetula is also microanimalivorous, feeding mainly on small mollusks ( Pais, 2019). The described trifurcated barbels are possibly structures associated with the search, selection, or manipulation of food.
In Chaco, this species was captured along with Ageneiosus militaris Valenciennes 1835 , Cetopsis gobioides Kner 1857 , Luciopimelodus pati (Valenciennes 1835) , Megaleporinus obtusidens (Valenciennes 1837) , Oxydoras kneri Bleeker 1862 , Pimelodus albicans (Valenciennes 1840) , Prochilodus lineatus (Valenciennes 1837) , Pseudohemiodon laticeps (Regan 1904) and Rhinodoras dorbignyi (Kner 1855) ; in Entre Ríos, with Ageneiosus inermis (Linnaeus 1766) , Lycengraulis grossidens (Agassiz 1829) and Rhinodoras dorbignyi ; in Buenos Aires, with Brochiloricaria chauliodon Isbrücker 1978 and Prochilodus lineatus .
The resulting distribution is extensive ( Fig. 2 View FIGURE 2 ), with records in the middle and lower reaches of the Paraná River and upper and lower sections of the Uruguay River, in Argentina, Brazil and Uruguay.
The category assigned to this species for IUCN is DD (Data Deficient) (ICMBio 2021). Due to the updated and wider geographical distribution that was found herein, this category could be changed to LC (Least Concern). Still, because of its rarity and the lack of knowledge about its biology, we prefer to maintain the current DD category.
The shortage of material in collections and publications may be more related to the habitat of the species than to its actual abundance; all new specimens were captured in deep environments of the main channel. In other large rivers of South America, the bottom sectors of the main channels (tens of meters deep) have been relatively poorly sampled; however, they have been revealed as habitat sometimes exclusive of a significant number of species, mainly Siluriformes and Gymnotiformes, adapted to an environment of very little or no light, turbulent waters, and high current speed ( Crampton 2007). According to Thomé-Souza & Chao (2004) in Negro River and Lasso & Morales-Betancourt (2022) in Orinoco River, these environments are used by many species (134 and 106 species, respectively). In the Paraná River in Argentina, there is a minimal history of this type of works; only a single bottom sampling campaign would have been carried out in 1975 with the ship “ARA Petrel” ( Calviño & Castello 2008). In particular, the specimens collected in the present work were captured with batteries of drifting, bottom nets, a method not used before in scientific fishing in the country.

| T |
Tavera, Department of Geology and Geophysics |
| MCP |
Pontificia Universidade Catolica do Rio Grande do Sul |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |