
Alvinocaris longirostris Kikuchi & Ohta, 1995
|
publication ID |
https://doi.org/ 10.5281/zenodo.168897 |
|
DOI |
https://doi.org/10.5281/zenodo.6270890 |
|
persistent identifier |
https://treatment.plazi.org/id/3A0287AC-FFEE-0260-FEBF-5456FB3DFA2D |
|
treatment provided by |
Plazi |
|
scientific name |
Alvinocaris longirostris Kikuchi & Ohta, 1995 |
| status |
|
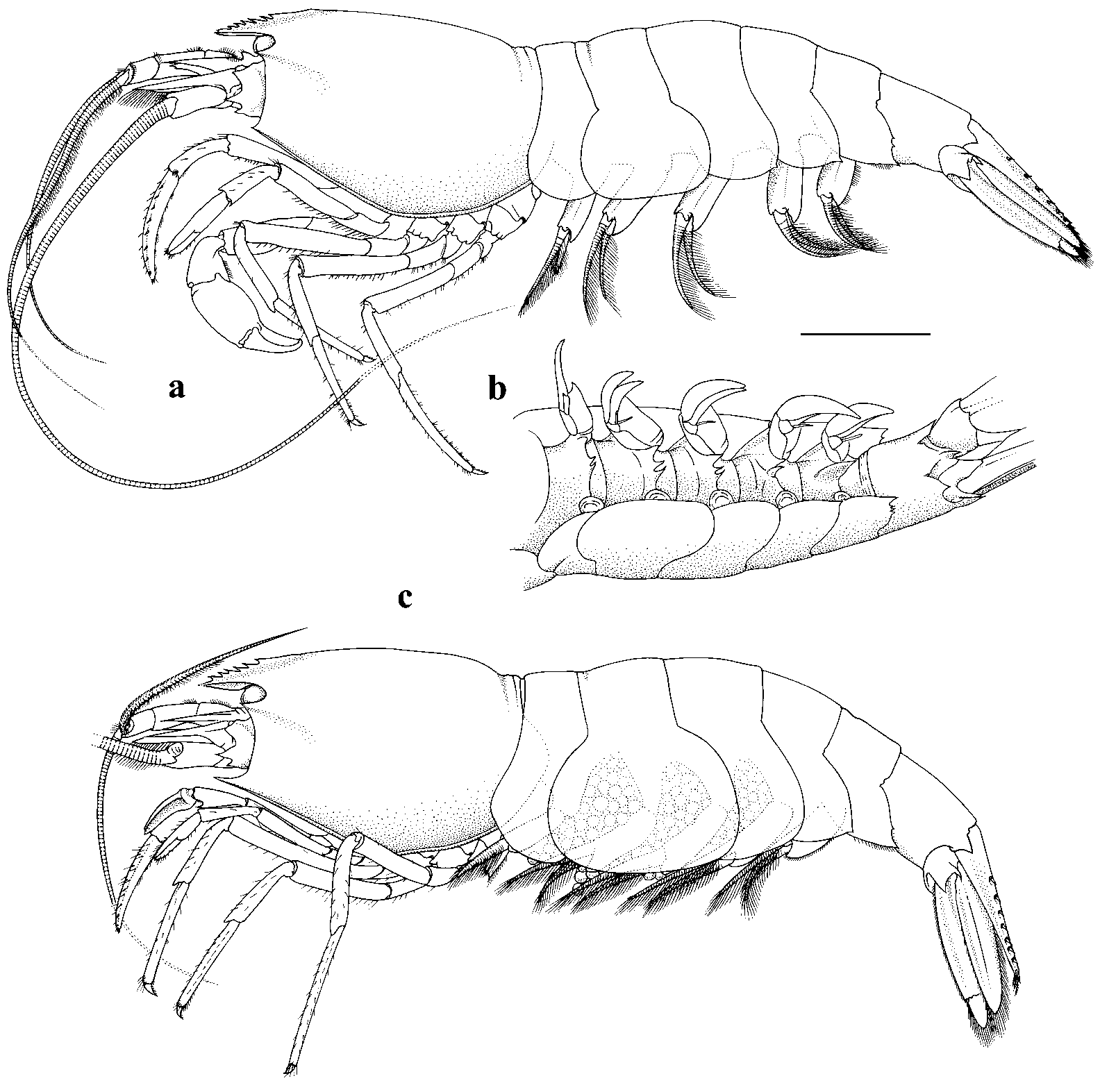
Alvinocaris longirostris Kikuchi & Ohta, 1995 View in CoL ( Fig. 5 View FIGURE 5 , 6 View FIGURE 6 )
Alvinocaris longirostris Kikuchi & Ohta, 1995: 772 View in CoL , Figs. 1 View FIGURE 1 –7. Alvinocaris View in CoL sp. B. — Webber & Bruce, 2002: 6, fig.
Material examined: —1 ɗ, 1 Ψ, R.V. Tangaroa stn X553, Brothers Caldera, 2 February 1996, 34° 52.77–53.25´S 179° 4.33–4.59´E, 1,335–1,490 m, rock dredge [ NIWA 3274 (ɗ), 3275 (Ψ)]; 1 ɗ, 5 Ψ, R.V. Tangaroa stn TAN0107/131, Brothers Caldera, 21 May 2001, 34° 52.58–52.28´S 179° 3.80–3.64´E, 1,370– 1,200 m, benthic sled [ NIWA 3263 (specimens 131, D–F), NIWA 3269 (52 specimens); MNZ CR. 9978 (specimen A), MNZ CR. 9988 (5 specimens)]; 1 Ψ, R.V. Tangaroa stn TAN0107/134 Brothers Caldera, 21 May 2001, 34° 52.87–52.76´S 179° 4.17–4.60´E, 1,518– 1,210 m, epibenthic sled [ NIWA 3264 (1 study specimen), NIWA 3270 (6 specimens); MNZ CR. 9985 (2 specimens)]; 3 Ψ, R.V. Tangaroa stn TAN0107/135, Brothers Caldera, 34° 52.89–52.87´S 179° 3.76–3.21´E, 1,346– 1,196 m, benthic sled [ NIWA 3265 (specimens A, C), NIWA 3271 (94 specimens); MNZ CR. 9979 (specimen B), MNZ CR. 9987 (5 specimens)]; 1 ɗ, 2 Ψ, R.V. Tangaroa stn TAN0107/136 Brothers Caldera, 21 May 2001, 34° 53.12–53.35´S 179° 4.49–5.09´E, 1,526– 1,197 m, benthic sled [ NIWA 3266 (specimens A, B), NIWA 3272 (80 specimens); MNZ CR. 9980 (specimen A), MNZ CR. 9986 (5 specimens)]; 1 Ψ, R.V. Tangaroa stn TAN0107/140, Brothers Caldera, 22 May 2001, 34° 51.69–51.46´S 179° 3.35–3.11´E, 1,850– 1,460 m, benthic sled [ NIWA 3267 (1 study specimen)]; 7 ɗ, 20 Ψ (2 ovig.), R.V. Tangaroa stn TAN0107/141, Brothers Caldera, 34° 52.96–52.69´S 179° 4.02–4.93´E, 1,538– 1,197 m, benthic sled [ NIWA 3262 (illustrated specimen), NIWA 3268 (specimens 141vou, BU, Y, Z), NIWA 3273 (73 specimens); MNZ CR. 9981 (specimen V), MNZ CR. 9982 (specimen X), MNZ CR. 9983 (specimen W), MNZ CR. 9984 (10 specimens)].
Specimens listed above with a letter or similar identifier were measured and examined in detail; those lacking such a label were identified as A. longirostris but not measured. No specimens from the following station were examined in detail: R.V. Tangaroa stn TAN0107/130 Brothers Caldera, 21 May 2001, 34° 53.30–53.69´S 179° 4.00–3.91´E, 1,350– 1,197 m, benthic sled [ NIWA 3276 (24 specimens)].
Fortyone specimens of A. longirostris were measured ( NIWA 3263–3268) and a study specimen illustrated and dissected (female NIWA 3262, CL 12.1 mm; RL 15.2 mm (together giving total carapace length of 15.38 mm); CD 9.4 mm; TL approximately 58 mm). A range of sizes was chosen from the approximately 400 specimens collected (dismembered specimens preclude a precise count) including at least one from each station at which the species was found.
A. longirostris Kikuchi & Ohta, 1995 View in CoL was described from 21 female specimens. Among the 41 New Zealand specimens examined nine were males.
A. longirostris View in CoL ( Figs 5 View FIGURE 5 , 6 View FIGURE 6 ) from Brothers Caldera agrees with the description of the species by Kikuchi & Ohta (1995) (hereafter K& O) in all its major characters, as follows: Rostrum slightly curved upwards distally; reaching beyond distal margin of the antennal peduncle ( Figs 5 View FIGURE 5 , 6 View FIGURE 6 a). Rostral dorsal margin armed with large teeth ( Fig 1 View FIGURE 1 a in K& O); ventral margin with smaller teeth. Prominent antennal and pterygostomian spines; antennal groove running backwards obliquely from antennal spine. Strong median sternal spine between fifth pereopods ( Fig 1 View FIGURE 1 c in K& O).
Pleura of abdominal somites 3–5 with variable number of spines ( Fig. 5 View FIGURE 5 this paper; Fig 5 View FIGURE 5 , K& O). Telson with dorsolateral row of seven movable spines on each side; two pairs of spines on posterolateral corners; posterior margin with row of setae (see variation below for ranges of spines and setae in Brothers material).
Antennular peduncle ( Fig. 6 View FIGURE 6 a) of similar reach to antennal scale; stylocerite reaching mid length of middle segment. Antennal scale with strong distolateral spine; basal segment with strong ventral spine. Mandibular incisor process with subterminal tooth on distal margin ( Fig. 6 View FIGURE 6 b). Third maxilliped with three spines at tip; carpus with acute spine distally; exopod reduced to fingerlike projection. Dense patch of setae on carpus of P1 with two larger spines and row of smaller spines, adjacent to it. P2 ischium with single stout spine on ventromesial face; P3–P5 of similar length with meri progressively shorter, propodi progressively longer from P3–P5; single row of spines on flexor surfaces of dactyls ( Fig. 6 View FIGURE 6 e). Pleopods 1–5 with a slender appendix interna, appendix masculina with about six slender setae. Uropod rami slightly longer than telson; movable spine laterally on exopod.
Variation ( A. longirostris , New Zealand). The format followed here is similar to that for A. niwa .
Carapace: The number of carapace dorsal teeth ranges 10–16 (n = 41) including 6–11 rostral; 3–7 carapace. The rostral ventral teeth range 5–14 (n = 41); 24 (58%) of these have 7–9 teeth while one has 12 and one 14.
Abdominal pleura: On abdominal somites 3–5, the number of spines on pleura are as follows (sample number in brackets):
The number of spines on AP3–AP5 varies considerably. Spines are usually present on AP3 with the majority of AP3 pleura having 3–5 spines, but the number is very variable and wide ranging. Only 12 (n = 74) shrimps were found with the same number of spines on both left and right pleura of AP3; no AP4 pleura examined lacked spines. AP4 usually has a single, more prominent posterolateral spine but in five pleura spination was continuous and undifferentiated around the posterolateral curve; AP5 almost invariably has a major posterolateral spine (a single pleuron lacks spines altogether), 74% of AP5 pleura have either three or four spines usually above the major spine on the posterior margin of the pleuron although a single specimen has spines (three) on the lateral border anterior to the major spine (for illustrations of variation in pleural armature of Alvinocaris longirostris , see K& O, Fig. 5 View FIGURE 5 , page 777).
Pereopod spines: On the ischia and meri of P2–5, the numbers of ventrolateral spines are as follows:
The number of ischial and meral spines on P2–5 varies throughout the 41 shrimps examined but least on the ischium and merus of P2. The two counts of zero spines on the P2 ischium and one spine on the merus, occur on a single animal (i.e. both conditions are paired); the ischia of AP3 and AP4 usually have two spines although variation from this number is 43% in AP4; the meri of AP3 and AP4 usually have three spines but variation from this number is 39% in AP3 and 65% in AP4; the merus of AP5 is the least predictable with nearly as many having one spine as two spines while 17% lack spines altogether.
Telson; the number of dorsolateral spines ranges 5–9 (n = 59) on both left and right sides; 12 (n = 58) individuals have a different number on each side, the difference ranging up to two. The number of posterolateral corner spines is almost always two but on three posterolateral corners (n = 59) there is one spine and on two there are three spines.
In common with A. niwa the telson posterior margin varies from distinctly bilaterally convex with a ‘v’–‘u’ shaped median indentation of varying size, to a single, convex curve with no indentation; the indentation is sometimes unevenly shaped and not always centred on the telson midline. Also like A. niwa , a tiny spinule is present in the apex of the indentation in most specimens and is also sometimes present at or near the centre in specimens with a single convex margin.
The number of telson posterior setae is about 0.5 that of A. niwa but also varies, as does their distribution. The total number of setae ranges 7–14 (n = 30). In all 30 specimens complete enough for examination a central gap divides setae into left and right series ranging 4–7 (left side), 3–8 (right side). Only nine (n = 30) specimens have the same number on left and right sides and, like A. niwa , a few posterior setae are sometimes submarginal.
Correlation analysis: As in A. niwa there is a strong positive correlation between CL and RL (r = 0.87, n = 40, p<0.001). However, there was no significant correlation between CL and any of the other characters analysed, viz. carapace dorsal spines (r = 0.20, n = 40, p>0.2), right P3 merus (r = 0.03, n = 33, p>0.5), right abdominal pleuron 5 (r = 0.08, n = 40, p>0.5), right telson dorsolateral spines (r = 0.14, n = 29, p>0.2) and telson posterior setae (r = 0.23, n = 28, p>0.2). This lack of correlation holds for both females and males, although the male sample is very small with data for less than ten counts from each character available for analysis.
Remarks: Alvinocaris longirostris was described from three stations on the Iheya Ridge of the Okinawa Trough at 1350–1410 m depth (K& O). The New Zealand A. longirostris were collected at eight stations at close proximity on the Brothers Caldera between 1,850– 1,197 m. Each tow on the Brothers covered a depth range from 150 to 390 m resulting in considerable overlap in collection depths. These depths encompass those sampled in the Okinawa Trough. Stn 140 (1850– 1460 m) indicates that A. longirostris also live at greater depths north of New Zealand than reported for the Okinawa Trough and stn 130 (1350– 1197 m) suggests they inhabit shallower depths there as well.
The only difference between the New Zealand specimens of A. longirostris and the original description of the species (K& O) which does not reconcile is the ratio of rostrum length: carapace length. In the study female the rostrum ( Fig. 5 View FIGURE 5 , 6 View FIGURE 6 a) is 1.26 times the length of the carapace. In the similar sized illustrated female holotype (CL 12.5 mm) the rostrum is about 1.2 times the length of the carapace (rostrum measured on K& O, Fig. 1 View FIGURE 1 , page 772). The RL:CL ratio in the holotype is thus about 0.83. The following table separates the RL:CL ratios of the New Zealand specimens into four divisions to indicate the range of shrimp sizes found with these ratios:
Ratios of rostrum length: carapace length in New Zealand A. longirostris (n = 40) A wide range of carapace lengths is associated with each division of the ratios and there is considerable overlap of the size classes, with one another. Despite this, with a carapace length fractionally greater than the New Zealand study specimen, the RL:CL ratio in the holotype (0.83) falls into the 5.80–10.75 mm CL size class, outside the range of ratios of New Zealand specimens of that size. However, the RL:CL ratio is clearly very variable which may render the sample size (40) limited with respect to this comparison. This, and the rostral measurement being taken from an illustration do not necessarily preclude the unusual ratio found in the holotype from falling within the range of variation of this character.
Other, minor differences between the New Zealand and Japan material include setae not reported in the original description; the presence of a single distolateral plumose seta on the proximal segment of the mandibular palp ( Fig. 6 View FIGURE 6 b); on the third maxilla ( Fig. 6 View FIGURE 6 c) and first maxilliped ( Fig. 6 View FIGURE 6 d) of the New Zealand study specimen, a small number of very plumose setae are present adjacent to the mesial margin of the expanded exopods and are present on both anterior and posterior faces. New Zealand A. longirostris otherwise key out similarly to their Japanese counterparts in the key to Alvinocaris species of Kikuchi & Hashimoto (2000).
The size of the first pereopod (cheliped) is dimorphic, as it is in A. niwa , with the palm inflated in males and similar in length to the fingers while it is hardly inflated in females and about half the length of the fingers. The male second pleopod showing the appendix interna and appendix masculina, with about six slender terminal and subterminal setae, is illustrated in Fig. 6 View FIGURE 6 f.
General comment: The morphological features usually employed to characterise species of Alvinocaris (e.g. A. lusca Williams & Chace, 1982 ; A. williamsi Shank & Martin, 2003 ) are very variable in the two species from northern New Zealand. Correlation analyses also show that, except for a strong positive correlation between carapace length (size) and rostrum length in both A. niwa and A. longirostris , and between size and the number of telson setae in A. niwa , most of the other characters quantified do not relate to size. These results indicate that numbers of teeth, spines and setae are determined genetically and cannot be interpreted as growth related. Authors such as K& O (describing A. longirostris ) have also recorded variation. However, descriptions of some species are, understandably, based on limited material (e.g. A. markensis and A. muricola Williams, 1988 , each described from three specimens). The level of variation in A. niwa and A. longirostris suggests that recognised species of Alvinocaris could be subject to future revision. It also indicates that molecular characters, already recorded for several alvinocaridid species by Shank et al. (1999) and Shank & Martin (2003), may be particularly useful in the taxonomy of these shrimps, which should be collected and preserved with this in mind.


| NIWA |
National Institute of Water and Atmospheric Research |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Alvinocaris longirostris Kikuchi & Ohta, 1995
| Webber, W. Richard 2004 |
Alvinocaris longirostris
| Webber 2002: 6 |
| Kikuchi 1995: 772 |