
Serpula hartmanae Reish, 1968
|
publication ID |
https://doi.org/10.11646/zootaxa.2848.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/396387E7-5F01-E044-FF50-FB91FBB2FBCB |
|
treatment provided by |
Felipe (2021-08-23 12:03:19, last updated by Plazi 2023-11-04 12:39:19) |
|
scientific name |
Serpula hartmanae Reish, 1968 |
| status |
|
Figs 28–32 View FIGURE 28 View FIGURE 29 View FIGURE 30 View FIGURE 31 View FIGURE 32 , Table 4
Serpula hartmanae Reish, 1968: 228–229 , figs 5, 11–16 [Type locality: Marshall Islands ( Fig. 28 View FIGURE 28 ), Bikini Atoll, Enyu Island]; Imajima & ten Hove 1984: 36–38, figs 1a–d [ Marshall Islands]; Imajima & ten Hove 1986: 2 [ Solomon Islands, Gilbert Islands].
Serpula sp. : Hartman 1954: 641 [northern Marshall Islands, Eniwetok Atoll].
Eastern Mediterranean
No previous records, but see Remarks.
Suez Canal
No previous records.
Gulf of Suez and Gulf of Aqaba
Serpula concharum not Langerhans, sensu Amoureux et al. 1978: 143, fig. 11, as S. c. [part; Gulf of Aqaba, Elat, Stn 6846, underside of a buoy, legit L. Fishelson, 18.IX.1970, on dead corals, 2 specs, synonymised herein ( Fig. 29 View FIGURE 29 ).
Red Sea proper–Indo-West-Pacific (excluding citations from Gulf of Suez and Gulf of Aqaba already given above) Serpula concharum not Langerhans, sensu Imajima 1982: 38–39, figs 1a–i [fide Imajima & ten Hove (1984),
Micronesia, Palau and Yap Islands].
Material examined. Locations adjacent to the Suez Canal, Mediterranean side: Questionable (somewhat similar) material examined (see below).
Suez Canal material reported herein: Great Bitter Lake “Yellow Fleet” Biofouling Samples , January 13– 20, 1975: 4 subsamples, 4 specs, two specimens with normally developed opercula (larger, B&T-02, HUJ- Poly-4388 ( Figs 30 View FIGURE 30 , 31 View FIGURE 31 ); smaller, B&T-01, HUJ-Poly-4387 ( Fig. 32 View FIGURE 32 ); and two with small, regenerating opercula, B&T-03, HUJ-Poly-4390, respectively B&T-04, HUJ-Poly-4389) .
Locations adjacent to the Suez Canal, Red Sea side: Gulf of Suez: None.—Gulf of Aqaba: Israel, Elat, Serpula concharum not Langerhans, sensu Amoureux et al.: 1978: 143, fig. 11, as S. c. [part; Gulf of Aqaba, Elat, Stn 6846, underside of a buoy, legit L. Fishelson, 18.IX.1970, on dead corals, 2 specs ( Fig. 29 View FIGURE 29 )— Egypt, Taba Beach, under rock, 11.V.1988, 3 questionable specs, see remarks below.
Indo-West-Pacific: Marshall Islands, Bikini Atoll, Ocean Side of Enyu Island, legit, det. D. Reish 6.IX.1956, holotype USNM 38400, paratype 38401 ( Fig. 28 View FIGURE 28 ).— Solomon Islands, Mamara Point and Komimbo Bay, British Royal Society Expedition to the Solomon Islands, 1965, attached to coral on reef platform, legit, det. P.E. Gibbs, BM (NH) 1970.833–835, 3 specs.— Palau & Yap Islands, off Arumonogui, Japanese Expedition to the Palau and Yap Islands, VII.1980, M. Imajima (1982) det. S. concharum , redet. Imajima & ten Hove (1984) S. hartmanae, ZMA V.Pol. 3452, 1 spec. (of the 15 specs collected), tube.
Suez Canal depth and substrates: On Brachidontes pharaonis bivalves; on a barnacle; on a bryozoan.
Distribution. Red Sea: Gulf of Aqaba; Indo-West Pacific:? Seychelles;? India, Micronesia.
Description, based on the largest individual from the Great Bitter Lake with the most differentiated operculum, B&T-02 ( Figs 30 View FIGURE 30 , 31 View FIGURE 31 , Table 3). Length, 26.5 mm plus (some posterior chaetigers lacking); ca. 120 chaetigers (7 thoracic); operculum with 15 blunt radii without tubercles, apical grooves nearly reaching the centre, surface, concave, cup-like, depth ca. ½ the length of the vesicle. Profile bell-shaped with a waist, basal bulb slightly expanded, distal diameter 3.5 times diameter of waist which is positioned 41 % from the base ( Fig. 30D View FIGURE 30 ); length of external grooves 42.5 %. Peduncle with a slight but distinct ventral asymmetrical boss proximal to the marked constriction (more marked in glycerine mount than in alcohol, Figs 30D, F View FIGURE 30 , respectively). Number of branchial radioles per lobe, ca. 11.
Hyaline bodies in the branchial radioles present in all four Suez Canal specimens ( Figs 31F, G View FIGURE 31 and 32E View FIGURE 32 ) and in a Palau Island individual as well. Apron present.
Meristic characters are detailed for each of the four Suez Canal specimens in Table 3: Length, 12.7–26.5 + mm; width, 0.8–1.2 mm; number of chaetigers, 80–121+ (7 thoracic), with opercula radii numbering 12–16 (mature opercula with 12 and 15 radii and the regenerating opercula with 15 and 16 radii). The number of radii in Amoureux et al. ’s (1978) synonymised nominal Serpula concharum specimen was also 12; the paratype has 18 radii ( Fig. 28A View FIGURE 28 ). The number of branchial radioles in the lobe ~8–11 (~8, 10, 10, 11), respectively (largest individual underlined). Length of external grooves, ~37–44 %; the maximum opercular diameter 1.8–3.5 times that of waist; waist positioned 28.9–50 % from the base. Contour bell-shaped, with waist less developed in small regenerating opercula. Characteristic asymmetrical peduncular boss present in B&T-02, much less distinct in B&T-01, not present in the two regenerating opercula.
Collar chaetae: Bayonet chaetae with 2 conical teeth; in some of the chaetae, a small (fine), very easily overlooked tooth, positioned behind and between the two large teeth and the blade ( Figs 28 E View FIGURE 28 1 View FIGURE 1 , 29I, J View FIGURE 29 ; 31A View FIGURE 31 , second chaeta from left; see also Fig. 32F View FIGURE 32 ). Collar capillary chaetae present. Other thoracic chaetae “limbate” and capillary chaetae ( Fig. 31B View FIGURE 31 ). Thoracic uncini saw-shaped, F+5 ( Figs 29M View FIGURE 29 , 31D View FIGURE 31 ), (F+4, three smaller individuals). Anterior abdominal uncini saw-shaped, F+4, two individuals; both F+4 and F+3 uncini in one individual; F+ 3 in one individual ( Figs 28G View FIGURE 28 , 31E View FIGURE 31 ). Abdominal uncini becoming rasp-shaped posteriorly and increasing both in numbers of teeth in vertical row and in numbers of teeth in transversal row (e.g., F:2:3:4:4: plus several rows of smaller teeth towards apex, e.g., Fig. 29L View FIGURE 29 ). Abdominal chaetae asymmetric flat-trumpet chaetae with ca. 25 teeth ( Figs 28F View FIGURE 28 , 29K View FIGURE 29 ; 31C View FIGURE 31 , 32I View FIGURE 32 replaced by capillary chaetae posteriorly (B&T-02- capillary chaetae in last 17 abdominal chaetigers + a few missing chaetigers. Hyaline bodies were present in the branchial radioles; becoming infrequent towards the proximal part of the radiole, noted in all four individuals from the Suez Canal ( Figs 31F, G View FIGURE 31 , 32E View FIGURE 32 ).
Tube: White with 3 rounded longitudinal ridges on flat upper surface, trapezoidal in cross-section, with transversal ridges; in more anterior parts of the tube the 3 longitudinal ridges are less pronounced, tube appears less rugose ( Figs 28C, D View FIGURE 28 ; 29A–C View FIGURE 29 ; 30A, B View FIGURE 30 ; 32A, B View FIGURE 32 ). Thin granular hyaline surface overlay present.
1) For characterizing the opercula, the following contour types are given: A–Funnel shaped (rather straight walls), B–Bell-shaped without flare or waist, C–Bell-(trumpet)- shaped with flare, without waist, D–Bell-shaped with flare and waist, without markedly expanded basal “bulb”, E–Bell-shaped with flare and waist, with markedly expanded basal “bulb”,— 2) Constriction of peduncle at the base of the opercular vesicle,— 3) The asymmetric boss at the distal end of the opercular peduncle, characteristic for Serpula hartmanae Reish : +++ = very prominent asymmetric boss; ++ prominent (see Fig. 29F View FIGURE 29 ); + visible; – lacking,— 4) Dentition of the collar chaetae, designated as follows: 2L = 2 large conical teeth; 2L, 1m = 2 large conical teeth with 1 smaller median tooth positioned between the teeth and the blade ( Fig. 29J View FIGURE 29 ); 2L, 1M denotes 3 large conical teeth positioned as above; note that Serpula hartmanae bayonet chaetae lack proximal rasps or additional dentition,— 5) For radiolar hyaline bodies see Figs 31F, G View FIGURE 31 , 32E View FIGURE 32 . Brattström & Taasen specimen no. 2 (removed from Bryozoa [ Figs 30–31 View FIGURE 30 View FIGURE 31 ]),— 6) Missing several pygidial segments ( Fig. 30E View FIGURE 30 ),— 7) Figs 30D, F View FIGURE 30 ,— 8 View FIGURE 8 ) Fig. 30C View FIGURE 30 ,— 9 View FIGURE 9 ) Figs 30D– G View FIGURE 30 ,— 10 View FIGURE 10 ) Fig. 30G View FIGURE 30 ,— 11 View FIGURE 11 ) 1–2 collar chaetae with small median tooth ( Fig. 31A View FIGURE 31 , 2 View FIGURE 2 nd chaeta from left),— 12) Fig. 31D, F View FIGURE 31 denotes the sharply pointed anterior tooth in Serpula uncini, here seen in profile view,— 13) Fig. 31E View FIGURE 31 ,— 14 View FIGURE 14 ) see Figs 31F, G View FIGURE 31 ; thinning out of radiolar hyaline bodies towards proximal part (base of radiole).— 15) Fig. 30B View FIGURE 30 ,— 16 View FIGURE 16 ) Abbreviations, “tu/t”, thoracic uncini in torus; “ac”, abdominal chaetae; “au/t” abdominal uncini per torus; “caps”, capillary chaetae in abdomen; number of uncini in thoracic torus, 49; number of abdominal chaetae in fascicle, 13; number of abdominal uncini in torus, 74; last 17 chaetigers with capillary chaetae. Brattström & Taasen specimen no. 1 (removed from Mollusca [ Fig. 32 View FIGURE 32 ]),— 17) Specimen lost after photographing,— 18) Counted on digital photo,— 19) Small operculum, not fully developed and not fully chitinised, without waist ( Fig. 32D View FIGURE 32 ),— 20) Fig. 32C View FIGURE 32 ,— 21 View FIGURE 21 ) Peduncular boss barely developed ( Fig. 32D View FIGURE 32 ),— 22) Fig. 32F View FIGURE 32 , frontal view, left chaeta, shows 2 teeth (intermediate area is not in focus), right chaeta clearly shows 2 large teeth and small median tooth between the large teeth and the blade,— 23) Fig. 32 G View FIGURE 32 ,— 24 View FIGURE 24 ) Fig. 32H View FIGURE 32 ,— 25 View FIGURE 25 ) Hyaline bodies present,— 26) Fig. 32A, B View FIGURE 32 . Brattström & Taasen specimen no. 4 (removed from Mollusca; specimen with regenerating branchial crown and operculum): 27) Branchial crown of small size,— 28) Constriction present, not marked, not chitinised,— 29) Basal boss not developed,— 30) Most of the bayonet chaetae with 2 pointed teeth, somewhat abraded; the most basal chaeta appears to have an intermediate tooth between these teeth and the blade,— 31) In one torus, dorsal uncini F+5, ventral uncini F+4. Brattström & Taasen specimen no. 3, removed from Cirripedia, specimen with regenerating branchial crown and operculum,— 32) Minute intermediate tooth present.— 33) The number of transverse rows increases posteriorly; frontally, the uncini are rounded, thus; only some of the teeth were in focus; mau F+5, seemingly saw-shaped; more posterior uncini F+7/3–4 sawto-rasp-shaped (compiled from two foci). Red Sea specimen TAU-NS-6188 ( Amoureux et al. 1978: 143) ( Fig. 29 View FIGURE 29 ): 34) Posterior abdominal part missing,— 35) Fig. 29E View FIGURE 29 — 36) Fig. 29D View FIGURE 29 ,— 37) Single unbroken chaeta, dissected from base of fascicle ( Figs 29I, J View FIGURE 29 )— 38) Fig. 29M View FIGURE 29 lateral view, 39) Fig. 29L View FIGURE 29 , — 40) Lateral longitudinal ridges higher than median ridge ( Figs 29A–C View FIGURE 29 ). Paratype from Marshall Islands, USNM 38401: 41) Returned to USNM before the difficult-to-observe minute middle posterior tooth was noted in other specimens (the chaetae are abraded but the left-most chaeta seems to show middle posterior tooth).— 42) Thoracic uncini from enlarged fig. 15 of Reish (1968),— 43) Returned to USNM prior to studying character,— 44) Tube with granular layer, covering ridges. IWP Palau, ZMA V.Pol. 3452 ( Imajima & ten Hove 1984: 36–38, figs 1a–d). IWP Seychelles, ZMA V.Pol. 4353, S. cf. hartmanae : 45) Very abraded chaetae with 2-teeth; figure of basal chaeta seems to show tip of small medial-tooth.
Remarks. Narrowing down the identification of our Suez Canal material of the genus Serpula , we considered the species neighbouring the Canal at both sides (and further away as well); see extensive discussion of this approach under Protula . The name Serpula concharum has erroneously been given to Indo- West-Pacific taxa, e.g., Amoureux et al. (1978, see above). Identifying “ Serpula concharum ”- like taxa with a similar opercular contour and a low number of blunt marginal radii from the Suez Canal area, we can discount both Mediterranean taxa of this nominal species (see remarks on S. concharum , above) by the fact that they lack the main diagnostic characters for S. hartmanae : The asymmetric ventral peduncular boss (swelling) proximal to the constriction of the operculum, termed by Reish (1968) “2 knobs”, and a granular surface overlay on the white tube (ten Hove 1994). However, in the Solomon Island specimens of P.E. Gibbs (1971), a sketch of spec. no. 833 showed a prominent asymmetric boss just below the opercular constriction while that of no. 835 was not prominent (notes, Ben-Eliahu, 1986), an indication as to the variability of this character.
Considering Serpula taxa distributed in the Mediterranean, we excluded S. cavernicola Fassari & Mòllica, 1991 , S. israelitica Amoureux, 1976 and S. vermicularis , all with more (40+) radii than our present material. It should be noted that we consider S. vermicularis strictly as an Atlantic / Mediterranean taxon, not cosmopolitan, cf. ten Hove & Jansen-Jacobs (1984). Serpula lobiancoi Rioja, 1917 has a pronouncedly zygomorphic operculum.
In considering described Indo-West-Pacific (IWP) taxa, we excluded several IWP taxa with more radii such as Serpula indica Parab & Gaikwad, 1989 (maybe including the very similar S. nudiradiata Pillai, 2009 , see further down), or S. jukesii , and taxa with a similar number of radii but with more teeth on the collar chaetae such as S. oshimae Imajima & ten Hove, 1984 that also has a predominantly orange tube and 7–9 thoracic chaetigers ( Imajima 1978, as S. cf. kaempferi) or S. vittata Augener, 1914 (syn. S. palauensis Imajima, 1982 ) that has Hydroides elegans -like collar chaetae and a tube dotted with brownish speckles. Another Indo-West-Pacific species, S. rubens Straughan, 1967b (p. 209–211, fig. 4a–i), with a similar opercular contour and number of radii (16–18), and collar chaetae with a large and several small teeth, is a much smaller species (to 8 mm in length), with fewer pairs of radioles (a maximum of 7 pairs), and with a variable number of thoracic segments, 8–11 ( Imajima & ten Hove 1984: 38).
The Suez Canal material comprised only 4 individuals and, regrettably, two of them, B&T-03 and -04, had regenerating opercula. Revising Serpula hartmanae parameters from Imajima (1982, as S. concharum ) and Imajima & ten Hove (1984) provides a reported length of up to 24 mm, with up to 105 chaetigers; thorax with typical constant seven chaetigers; 15–16 radioles, and 11–25 blunt opercular radii. Meristic differences between their S. hartmanae material and that of Reish (1968) appeared to these authors insufficient for taxonomic distinction ( Imajima & ten Hove 1984: 38). Ten Hove (1994) reported Seychelles material with an even greater number of opercular radii (24–35) as Serpula cf. hartmanae due to the presence of an asymmetric peduncular boss (“peculiar swelling”) and a granular tube surface. Moreover, some of the Seychelles specimens’ opercular grooves were wavy, not smooth, thus it might well be a separate species. The Red Sea S. hartmanae individual had 12 opercular radii ( Fig. 29D View FIGURE 29 ).
As Table 3 shows, the largest individual from the Suez Canal (B&T-02) slightly exceeds the compiled length described for Serpula hartmanae . Its operculum shows an asymmetric peduncular boss ( Figs 30D–F View FIGURE 30 ), though less prominent than that of the Red Sea specimen ( Fig. 29E, F View FIGURE 29 ) which is similar in size to that of the paratype ( Fig. 28B View FIGURE 28 )—it is smaller than that figured for the holotype by Reish (1968, fig. 11). A Palau Island individual had an even larger boss than the paratype or the Red Sea individual ( Imajima & ten Hove 1984: 37, fig. 1a).
The number of chaetigers in these Suez Canal specimens is also slightly greater than that described for Serpula hartmanae , but that is also true for other S. hartmanae individuals, e.g., from Palau Island (material of Imajima & ten Hove 1984); that of the Red Sea specimen with a missing posterior abdomen was 67+. The number of radioles in the Suez Canal material was less than that described for Serpula hartmanae , while the Red Sea individual had 15 radioles in the non-opercular lobe, which conforms to the description.
In the several descriptions, differences in the chaetal dentition of the bayonet collar chaetae also emerge. Reish (1968) figured them with 2 teeth, rather abraded in the paratype (see Figs 28E View FIGURE 28 1 View FIGURE 1 , E 2 View FIGURE 2 ); ten Hove & Jansen- Jacobs (1984: 149, fig. 2q) similarly noted 2 teeth without accessory teeth, however, Imajima (1982) and Imajima & ten Hove (1984: 36) referred to “2–3 heavy conical teeth and a variable number of small accessory teeth at base.” The collar chaetae of one of their Palau specimens ( Imajima & ten Hove 1984) were abraded, except for a basal chaeta with 2 large conical teeth; whether or not there is an intermediate tooth positioned between these teeth and the blade could not be seen; there were no accessory teeth at the base. The Red Sea specimen had only one unbroken bayonet chaeta at the base of the fascicle; it had a distinct small median tooth positioned between the conical teeth and the blade ( Figs 29I, J View FIGURE 29 ) similar to some of the Suez Canal bayonet chaetae, described above ( Figs 31A View FIGURE 31 , 32F View FIGURE 32 )—the remaining collar chaetae were broken. In a Seychelles specimen (of Serpula cf. hartmanae ), the median tooth was larger than that in the Red Sea and Suez Canal chaetae ( Table 3).
Thoracic uncini: An enlargement of Reish’s (1968) figure of the thoracic uncini (fig. 15) shows a dentition of F+6. The thoracic uncinus given by Imajima (1982: 39, fig. 1f) was F+7 teeth, and Imajima & ten Hove (1984) reported F+7 thoracic uncini. However, a single torus from a Palau Island individual, showed several uncini of F+6 dentition, while most were F+5. The Red Sea specimen showed F+5 dentition ( Fig. 29M View FIGURE 29 ). As noted, the largest Suez Canal individual had F+5 uncini ( Fig. 31D View FIGURE 31 ), while the thoracic uncini of the three others was F+4, a lower number than previously described for Serpula hartmanae .
Abdominal uncini: Reish (1968) did not specify dentition of the saw-shaped anterior abdominal uncini; however that of the paratype was F+4 ( Fig. 28G View FIGURE 28 ). The abdominal uncini described by Imajima (1982), and Imajima & ten Hove (1984: 36) are F+7 anterior and posterior uncini, with up to 4 horizontal rows of teeth in the transversal rows of the rasp-shaped posterior uncini. The Red Sea specimen’s anterior abdominal uncini were F+4 while the not quite posterior-most uncini were F+7/4.
All Suez Canal individuals show a trend of increasing the number of teeth in the vertical rows posteriorly, and the increase in number of teeth in the horizontal rows continued, detailed for some individuals in Table 3, which also shows the variability of dentition within a single torus (most individuals have a posterior dentition of F+7/4 [see “Treatment of the specimens in “Material and Methods” section]). For example, dentition of a posterior abdominal uncinus of specimen B&T–2 was F+7/4 (or 5) (detailed F:1:2:3:3:2:[4 or 5]:4), while in specimen B&T–4, in the same torus, the number of teeth were: F+8/4 (detailed F:1:1:1:2:3:4:4:2) in one uncinus, and F+7/4 (= F:1:1:2:2:3:4:3) in another (seen under compound microscope).
Abdominal chaetae: Abdominal chaetae cannot be properly described without SEM due to their transparent nature (ten Hove & Jansen-Jacobs 1984: 145); photographs ( Figs 28F View FIGURE 28 , 29K View FIGURE 29 , 31C View FIGURE 31 and 32I View FIGURE 32 ) show mostly the fibrils and not the transparent distal teeth; thus, counts of the teeth done under the compound microscope are not reliable.
Tube : The Micronesian material from Palau Island ( Imajima & ten Hove 1984: 36–38, figs 1c, d) appears more granular than that of the paratype, the Red Sea or the Suez Canal individuals, but all of the tubes appeared to have a thin granular layer. In the Gulf of Aqaba specimens from Taba Beach, the seeming lack of a granular surface on the tubes, despite a three-ridged cross-section, raises a question regarding the identification of the three specimens taken from under a rock (11.V.1988 [cited above]); the identification as Serpula hartmanae is supported by the low number of opercular radii, 17, 16 opercular radii, 11 radii in a juvenile with marked constriction present at the base of the operculum .
Meristic differences found by Imajima & ten Hove (1984) did not seem to them sufficient for taxonomic distinction. Similarly, the Suez Canal material, though showing some variation, should be attributed to Serpula hartmanae . Our results suggest that the degree of waistline in the profile of the operculum is a function of the opercular development (size) and this appears to be true also for the development of the characteristic asymmetric peduncular boss. It is not known whether the closely packed hyaline bodies observed in the branchial radioles have disappeared in some preserved specimens. What is clear is that variability in various characters in the taxon S. hartmanae needs still to be further explored, and particularly, variability in development of the peduncular boss (bosses), in structure of bayonet collar chaetae, dentition of the uncini as well as the effect of different environmental conditions on the formation and degree of tube granulation, and the thickness of the lateral keels. In this exploration, S. amplilobata Pillai, 2009 (pp.134– 135, figs 27a–k, 18A–E), recently described upon a single specimen only, should be taken into account as well. Apparently Pillai was not aware of the existence of S. hartmanae , since not referring at all to this taxon in his paper, while it is at least very similar—if not synonymous—with his newly described taxon regarding operculum, granular tube, collar chaetae, etc.
On reviewing the present paper, H. Zibrowius contacted H.A. ten Hove concerning a new finding by Zibrowius & Bitar that Serpula hartmanae is abundant on the Lebanese coast (Zibrowius & Bitar, in prep.). This finding should cause it to be classified as a Lessepsian migrant proper, sensu Por (see Introduction). The report galvanized us to examine Serpula material encrusted on recently obtained molluscs (samples of the Tel Aviv University mollusc collection), as well as our own collections from 1990, diagnosed as S. cf. concharum (by Ben-Eliahu & ten Hove 1992: 41). The specimens are clearly different from Serpula concharum Langerhans, 1880 s. str., from deeper waters. All 22 specimens surveyed have a three-keeled tube. However, in some tubes, the two dorso-lateral keels are much more prominent, more rounded, and more blunt than the thinner medial one (as in Serpula hartmanae [ Fig. 29B View FIGURE 29 ]), but in others (perhaps half of the tubes), the keels are all narrow and rather similar—very likely this is related to size / age, but that still needs verification. Tubes are mostly covered by a thin granular layer. Some of the opercula show a small but distinct ventral peduncular boss proximal to the marked constriction (n=3) as in Serpula hartmanae ( Figs. 29E, F View FIGURE 29 , 30D, F View FIGURE 30 , and 31D View FIGURE 31 ), but, in most specimens, the boss is not pronounced. The number of radii range between 10– 13.3 – 19 (n=22, SD 2.3). Perhaps the variability observed in the tube structure and the peduncular boss falls within the range of a single taxon, but we cannot presently be sufficiently certain to make a conclusive determination of that. We assume that Zibrowius & Bitar’s Lebanese record belongs to the same taxon, which, in the past, we had identified as S. cf. concharum . This raises the question whether this material belongs to a taxon indigenous to the Mediterranean, as forms with a 3-keeled tube have been mentioned under various names, the oldest probably being Serpula sulfurata Milne Edwards, 1836 . Alternatively, there is a reasonable similarity with our specimen from the Gulf of Aqaba ( Fig. 29 View FIGURE 29 ), presumably identical with S. hartmanae , which indeed might make the taxon yet another Lessepsian migrant. In view of the fact that the taxonomy of the genus Serpula is exceedingly problematic (see Remarks under Serpula jukesii , this paper), we have refrained from reaching a decision at the present time. A revision of S. concharum sensu lato, and all material of Indo-West-Pacific records of similar taxa, including S. hartmanae , is necessary (see remark on variability of this nominal taxon one paragraph above as well), but falls outside the scope of the present paper. Serpula cf. concharum , recently reported in shallow Egyptian waters (Alexandria) by El-Rashidy et al. (2009), is very possibly the same taxon as that found along the Israeli coast (see remarks under Serpula concharum , above).
Amoureux, L. (1976) Serpula (Paraserpula) israelitica, nouvelle espece de Serpulidae (Annelides Polychetes) et une petite collection annelidienne de la Mediterranee orientale. Bulletin du Museum d'Histoire Naturelle, Paris, (3) 404 (Zoologie 281), 1047 - 1059.
Amoureux, L., Rullier, F. & Fishelson, L. (1978) Systematique et ecologie d'annelides polychetes de la presqu'ile du Sinai. Israel Journal of Zoology, 27, 57 - 163.
Augener, H. (1914) Polychaeta II: Sedentaria. In: Michaelsen, W. & Hartmeyer, R. (Eds), Die Fauna Sudwest-Australiens. Ergebnisse der Hamburger sudwest-australischen Forschungsreise 1905, 5, 1, 170 pp.
Ben-Eliahu, M. N. & Hove, H. A. ten (1992) Serpulid tubeworms (Annelida: Polychaeta) - a recent expedition along the Mediterranean coast of Israel finds new population buildups of Lessepsian migrant species. Israel Journal of Zoology, 38 (1), 35 - 53.
El-Rashidy, H. H., Atta, M. M., Dorgham, M. M. & Hamdy, R. (2009) New northward polychaetes immigrants through the Suaz [sic] Canal. Proceedings of Coastal Zone 09, Boston, Massachusetts, July 19 to 23, 2009, 2 pp.
Fassari, G. & Mollica, E. (1991) Una Nuova Specie di Serpulidae (Annelida Polychaeta) di grotta. Animalia, Catania, 18, 261 - 267.
Gibbs, P. E. (1971) The Polychaete fauna of the Solomon Islands. Bulletin of the British Museum (Natural History) Zoology, 21 (5), 101 - 211.
Hartman, O. (1954) Marine Annelids from the northern Marshall Islands. Geological Survey Professional Paper, 260 - Q: 619 - 644.
Hove, H. A. ten & Jansen-Jacobs, M. J. (1984) A revision of the genus Crucigera (Polychaeta, Serpulidae); a proposed methodical approach to serpulids, with special reference to variation in Serpula and Hydroides. In: Hutchings, P. A. (Ed.). Proceedings of the First International Polychaete Conference, Sydney, 1983, The Linnean Society of New South Wales, Sydney, pp. 143 - 180.
Hove, H. A. ten (1994) 6.6. Serpulidae (Annelida, Polychaeta) from the Seychelles and Amirante Islands. In: Land, J. van der (Ed.). Oceanic Reefs of the Seychelles. Cruise Reports of the Netherlands Indian Ocean Program, 2. National Natural History Museum, Leiden, pp. 107 - 116.
Imajima, M. (1978) Serpulidae, Annelida, Polychaeta) collected from around Nii-jima and O-shima, Izu Islands. Memoirs of the National Science Museum, Tokyo, 11, 49 - 72.
Imajima, M. (1982) Serpulinae (polychaetous annelids) from the Palau and Yap Islands, Micronesia. Proceedings of the Japanese Society of Systematic Zoology, 23, 37 - 55.
Imajima, M. & Hove, H. A. ten (1984) Serpulinae (Annelida, Polychaeta) from the Truk Islands, Ponape and Majuro Atoll, with some other new Indo-West-Pacific records. Proceedings of the Japanese Society of Systematic Zoology, 27, 35 - 66.
Imajima, M. & Hove, H. A. ten (1986) Serpulinae (Annelida, Polychaeta) from Nauru, the Gilbert Islands (Kiribati) and the Solomon Islands. Proceedings of the Japanese Society of Systematic Zoology, 32, 1 - 16.
Langerhans, P. (1880) Die Wurmfauna von Madeira. III. Zeitschrift fur Wissenschaftliche Zoologie, Leipzig, 34, 87 - 143.
Parab, P. P. & Gaikwad, U. D. (1989) Occurrence and ecology of Serpula indica sp. nov. (Serpulidae - Polychaeta) from Ratnagiri coast. Journal of Ecobiology, 1, 223 - 232.
Pillai, T. G. (2009) Descriptions of new serpulid polychaetes from the Kimberleys of Australia and discussion of Australian and Indo-West Pacific species of Spirobranchus and superficially similar taxa. Records of the Australian Museum, 61, 93 - 199.
Reish, D. J. (1968) The polychaetous annelids of the Marshall Islands. Pacific Science, 22, 2, 208 - 231.
Rioja, E. (1917) Datos para el conocimiento de la fauna de Anelidos Poliquetos del Cantabrico. Trabajos del Museo Nacional de ciencias naturales de Madrid (Zoologia), 29, 1 - 111.
Straughan, D. (1967 b) Marine Serpulidae (Annelida: Polychaeta) of Eastern Queensland and New South Wales. Australian Journal of Zoology, 15, 201 - 261.
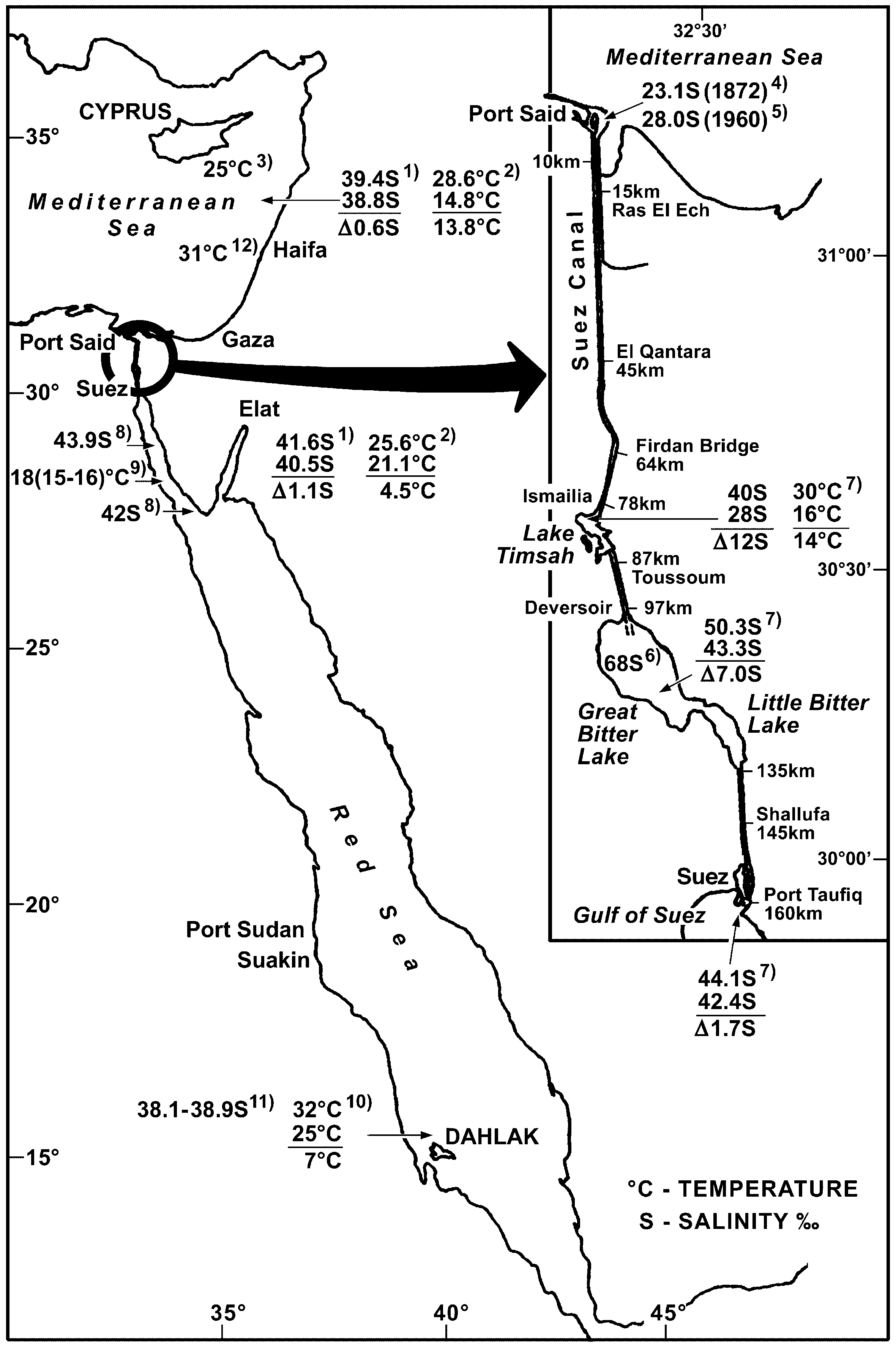
FIGURE 1. Map of Suez Canal showing salinity and temperature relations in the areas joined by the canal compiled from various sources. 1—Ben-Eliahu 1977: 70, Table 21, salinity ‰—seasonality, the extreme range of monthly means of sea surface salinity based on daily 00:08 recordings taken by the Nahariyya hydrographic station, northern Israel, 1968–1972 and the Elat hydrographic station, northern Gulf of Aqaba, 1962–1973, 2—Ben-Eliahu et al. 1988: 263, seasonality of sea surface temperature, Mediterranean coast of Israel and Elat, Gulf of Aqaba in °C (data from hydrographic stations listed above), 3—Por 1978: 118–119, fig. 32, southern Cyprus, summer surface isotherm 25°C; summer upwelling; 22°C, 4—Por 1978: 61, depressed salinity in Port Said due to Nile flood, 23.1‰ in autumn, 1872, 5—Ben-Tuvia 1970: 183, depressed surface salinity in Port Said due to Nile flood, autumn peak, IX & X.1960, 6—Thorson 1971: 842– 843, initial salinity at Great Bitter Lake, bottom, 8 m: 68–80‰; surface: 50–52‰, 7—Ghobashy & el-Komi 1981a: 169, 171, seasonality of Lake Timsah salinity and temperature between II.1977–I.1979; Ghobashy & el-Komi 1981b: 180, southern canal, seasonality of salinity and temperature at the Little Bitter Lake (Kabrit) and Suez between II.1977– I.1979, 8—Por 1972: 113–114. Gulf of Suez coast of Sinai, Ras el Missala and Ras es Sudr, 15 and 50 kms south of Suez, respectively, X.1970, Ras el Missalla along the shore, 44.25‰ and 25 m offshore 43.93‰; Ras es Sudr high VIII.1970, 44.25‰; X.1970, 41.69‰ and I.1971, 42‰, 9—Oren 1970: 226 reported 18°C temperature for the Gulf of Suez; however, Por 1972: 114, 1978: 83 noted even lower winter temperatures, particularly inshore, and found that the temperature decrease from south to north of Gulf of Suez corresponds with the depletion of the tropical fauna (i.e., of corals and associated taxa), from the south to the north of the Gulf, 10—Ben-Tuvia 1966: 255, mean monthly sea surface temperatures at Massawa, Eritrea, 11—Oren 1964: 12, table 3, III.1962, profile of Stn 7, surface to 110 m (low to high value, respectively) taken in the south Red Sea off Eritrea, off Entedebir and Dahlak Kebir Islands, 12—Brit 2000 (E. Spanier, pers. comm.): High peak temperature (9 m) prevailing off Haifa, northern Israel during August, 2000.

FIGURE 2. Biofouling on a ship that was trapped in the Suez Canal for 8 years when the canal was shut down due to the June 1967 war (adapted from Barracca & Thomas 1975). Scale: 1 m.
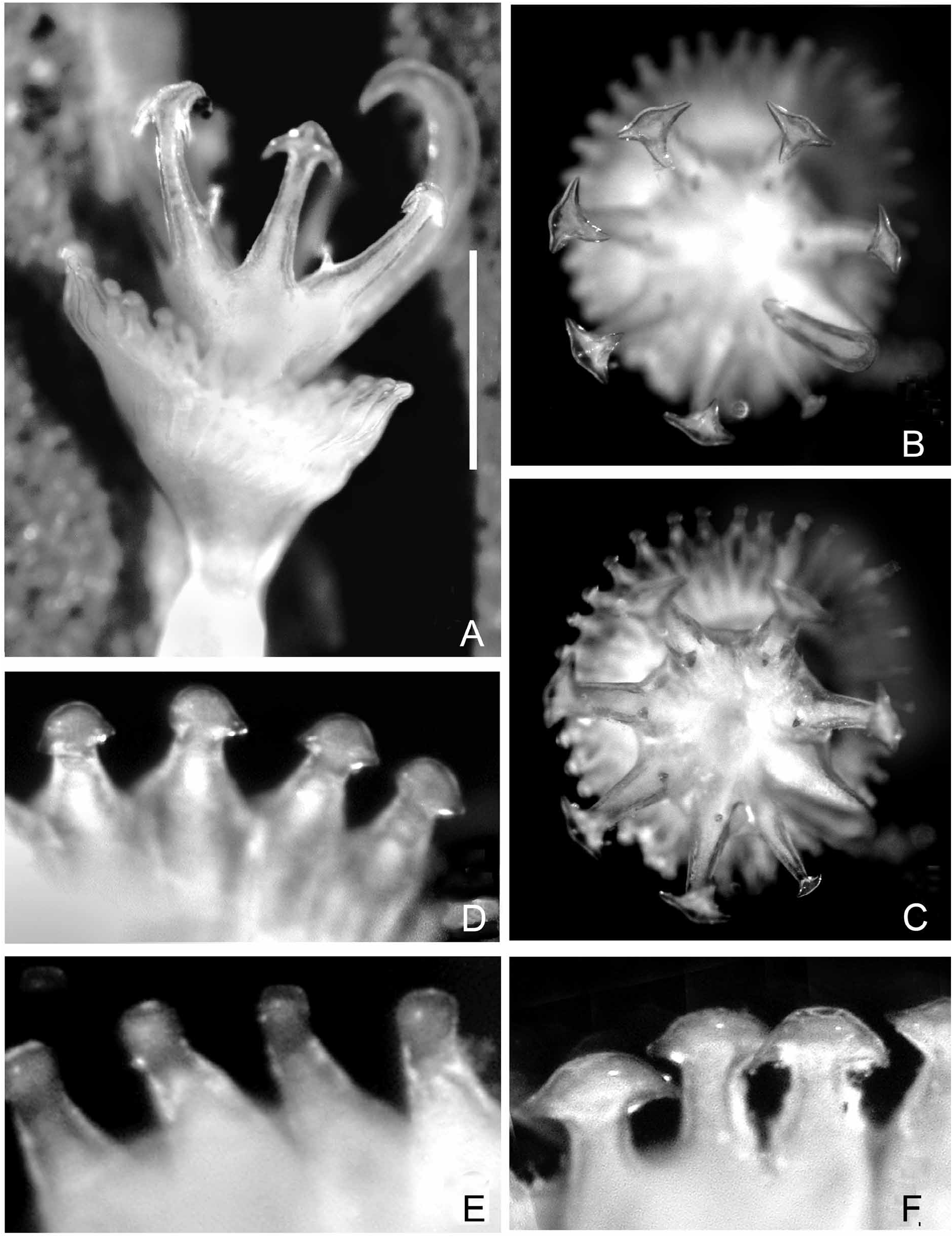
FIGURE 8. Hydroides heterocerus. Variability in forms of opercula from different regions: A–D. Operculum of specimen from the Great Bitter Lake aggregate removed by J.P. Taasen from the M/S “Münsterland” after its journey to the Hapag-Lloyd dry dock in Bremerhaven (18.VI.1975, 7 m). A—Lateral view, B–D—Apical views, B—Focus on tips of verticil spines, C—Focus on basal spinules of verticil, D—Enlargement of marginal teeth of funnel; the expansion of the tips of the marginal teeth in “D” is intermediate between “E” and “F,” E—“Bowling-pin-shaped” radii with blunt or even stubby tips, specimen from Little Bitter Lake opposite Kabrit (sample SLC 123), F—More expanded, “T”-shaped tips found in specimen from the Sudanese Red Sea (BM(NH) 1972.23, legit C. Crossland, 1904–1905). Lessepsian migrants with marginal radii like “D” or like “E”. Scale: 1 mm.
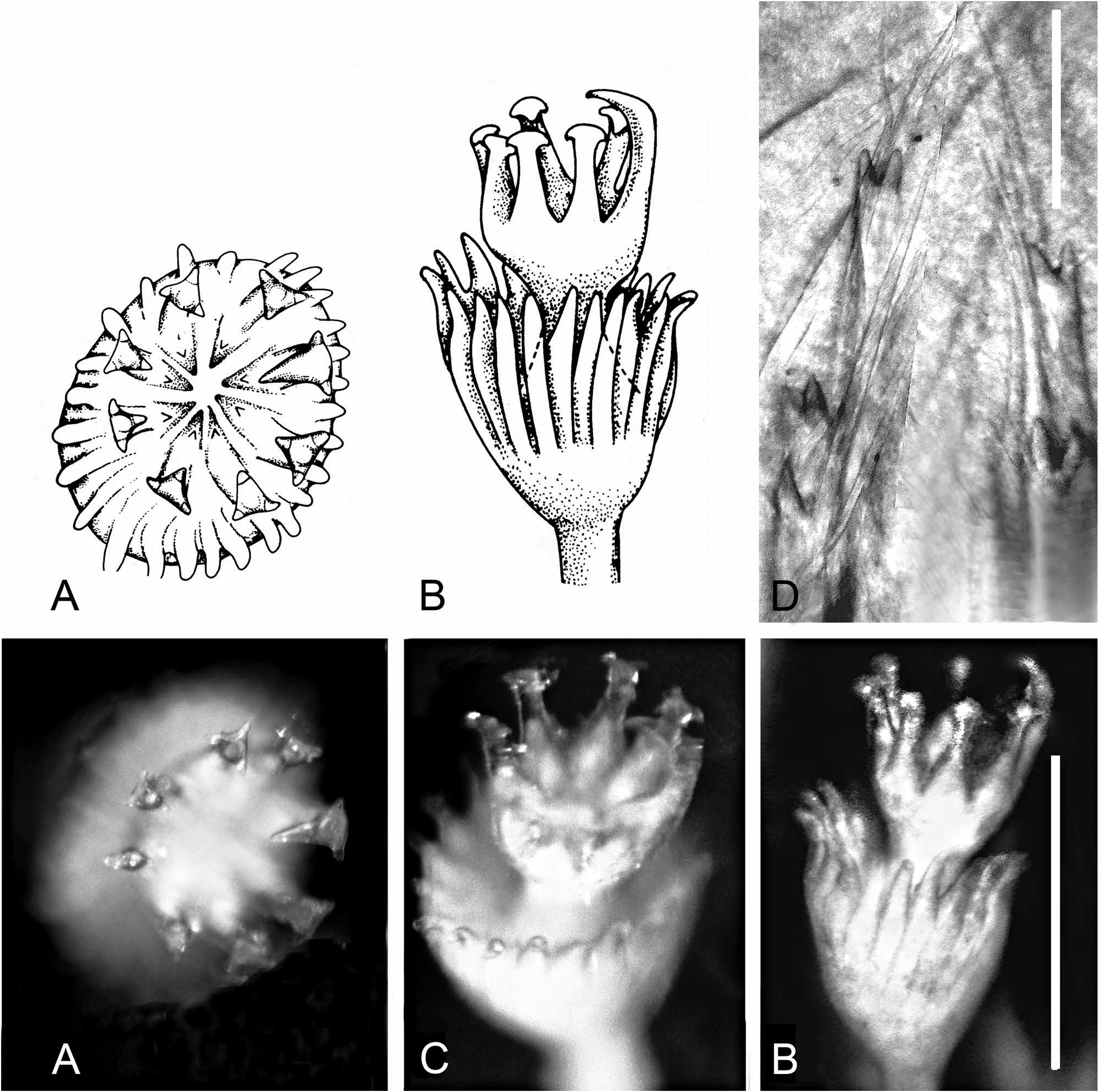
FIGURE 9. Operculum of Hydroides heterocerus juvenile form no. 1 (specimen from Cambridge Expedition mollusc deposited at the Natural History Museum in London, precise location unknown [See App. Table 2A]). A—Apical view, drawing and photograph, respectively, B—Profile view of drawing and photograph, C—Three-quarters view; note sharply pointed marginal teeth of funnel radii (in contrast with those in Fig. 8), D—Collar chaetae (magnification, <10 x). Scales: B—0.5 mm, D—100 µm.

FIGURE 10. Operculum of Hydroides heterocerus juvenile form no. 2 (specimen scraped from mollusc from the Great Bitter Lake [SLC 117, Y. Eytam, St. 4]). A, B—Funnel radii with some typical “anchor-shaped” (T-shaped) tips; some verticil spines approaching typical adult form, other spines deviating from the typical form, e.g., dorsal spine, upper right, bifid, aberrant. Scale: 100 µm.
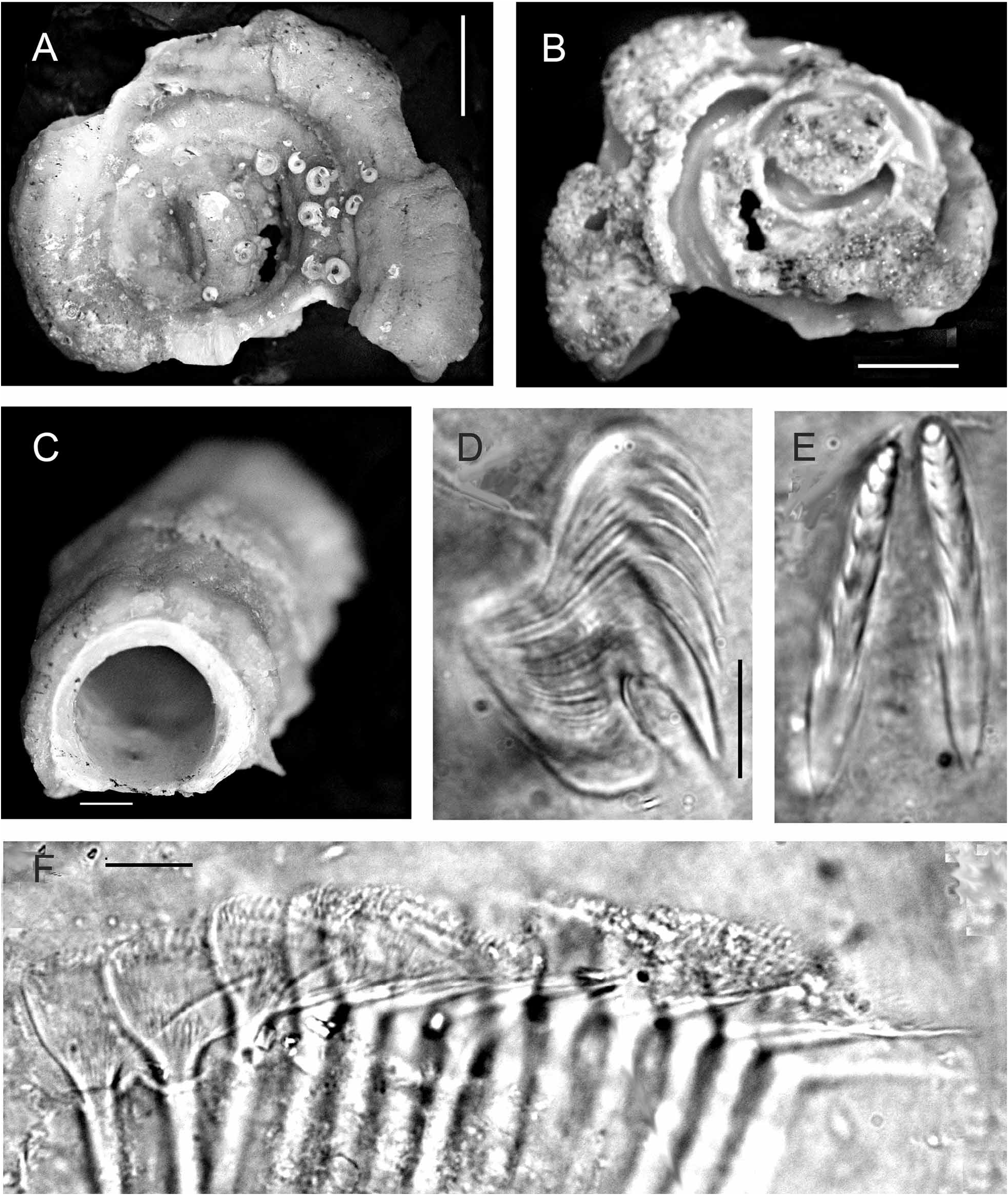
FIGURE 11. Large spiralled serpulid tube. The tube was present in the test-tube labelled "Serpula vermicularis" by F.A. Potts but should rather be attributed to Hydroides heterocerus (The nominal "Serpula vermicularis" specimen collected by the Cambridge Expedition in the Gulf of Suez that was described in detail by Potts 1928 was no longer present [CUZM]). The abdominal uncini and chaetae shown below were found on skin still attached to the lumen of the tube. A—Upper view of tube (note spirorbid tubes encrusted on outer surface of the tube), B—Ventral view of tube, C—3/4 frontal view of tube, D—Abdominal uncinus, lateral view, F+5, E—Same, frontal view, left, F+10, F—Abdominal chaetae. Scales: A, B—5 mm, C—1 mm, D, F—10 µm.
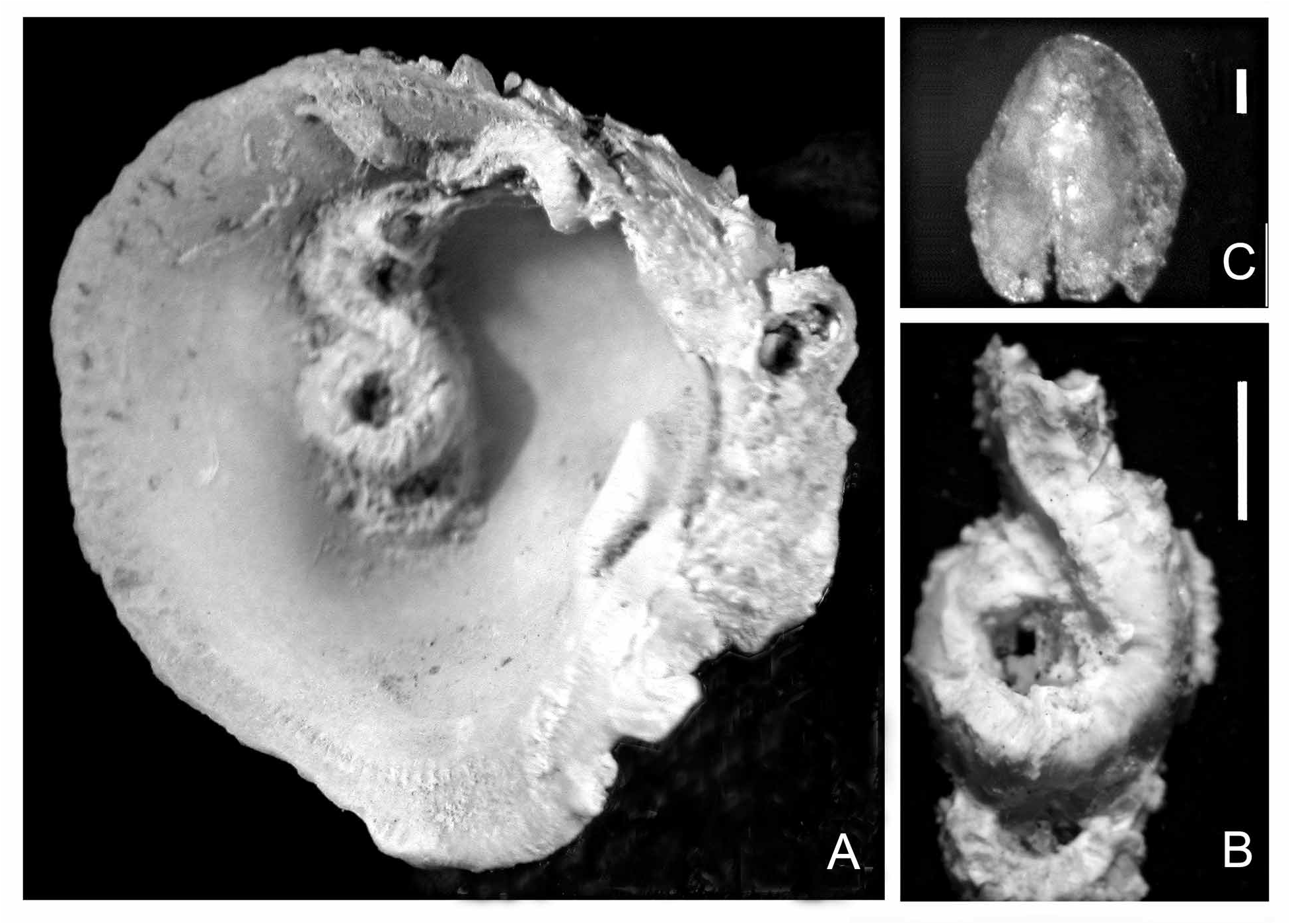
FIGURE 14. First record of Hydroides steinitzi in the Suez Canal from a taphonomic residue of the operculum. Operculum, collected in 1950, was within tube inside Chama asperella valve (Beets’ Stn 15, 10.4 m, see App. Table 2B). A— Tube in situ, B—Tube removed from valve; note upper surface somewhat encrusted with granular layer, C—Dried opercular residue (verticil) found within the tube had “lasted” 50 years (compare with those in Fig. 13). Scales: B—1 mm, C—100 µm.

FIGURE 16. The Hydroides “priscus” juvenile stage of Hydroides sp. / spp. A—From Lake Timsah, Isma’iliya, on bivalve Malvufundus normalis, Cambridge Expedition, 26.XI.1924, BM(NH) 1928.3.30.179, B—From sample Great Bitter Lake SLC 117 (see App. Table 2A, C, respectively). Scale: 100 µm.

FIGURE 21. Protula cf. palliata tubes from the Bitter Lake with distinctive sculpturing—simple growth striations, but with the rim of the striation seemingly eroded with a somewhat scalloped appearance. A–C—Tube fragment of Protula sp. on Chama asperella from Beets’ Stn 15, 10.4 m (Beets 1953): A—Inner view of Chama valve bearing tube of 3.6 mm diameter, B—Lumen of same tube fragment, diameter ca. 4 mm, C—Enlargement of A, D—Tube fragments of Protula specimen from “Yellow Fleet” encrustation (B&T-C-Pol 27-HUJ-Poly-1567), legit H. Brattström and J.P. Taasen, 1975, diameter 4.8 mm. Scales, 5 mm.
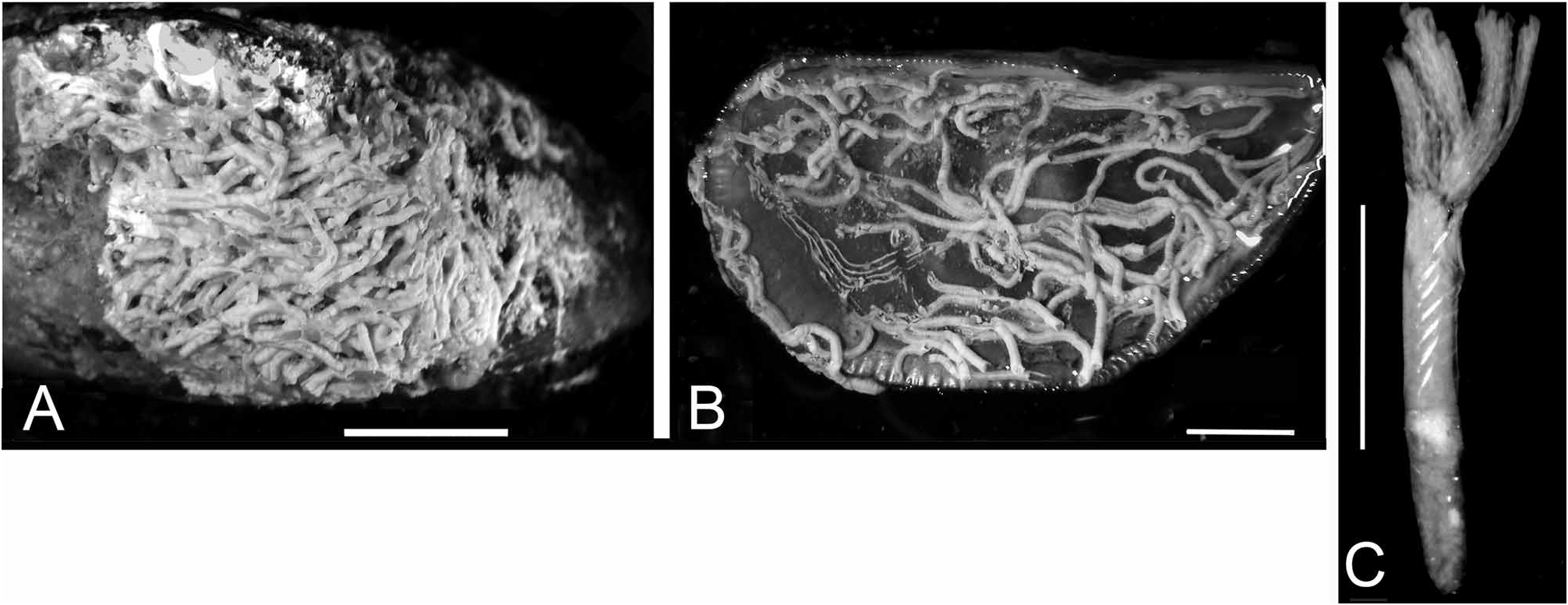
FIGURE 24. Salmacina incrustans encrusted on a Brachidontes pharaonis bivalve from the “Yellow Fleet”, Great Bitter Lake. A, B—Encrusting aggregation, outside and inside of a Brachidontes valve, respectively, C—Three-quarters view of worm (subsample Biv11). Scales: A, B–5 mm, C–1 mm.

FIGURE 25. Variability in fin and blade collar chaetae of Salmacina incrustans from the Great Bitter Lake sampled by H. Brattström and J.P. Taasen in January 1975. Note morphological variability of the fin structure in chaetae from the same individual. A1–3—Three collar chaetae (spec. no. 15), A1—Profile view, slightly darkened, A2—¾ view, A3—Frontal view of fin, B—Profile view of collar chaeta (spec. no. 8), C1–2—Frontal view of fin (spec. no. 14) with tips of blades of two chaetae to the left, D1–3—Three chaetae of spec. no. 10: D1—Frontal view of most distal fin, D2—Frontal view of fin positioned between the most distal and most proximal chaetae—the blade of these chaetae are below the focus level of the SEM and thus not visible in the micrograph; D3—¾ view of fin of most proximal chaeta, E—Frontal view of fin (specimen no. 16), F—Frontal view of fins (spec. no. 13; note the free (non-denticulate) space between the fin and the blades; the denticulate structure of the blades, G. G1—Frontal view of fin, G2—Lateral, almost profile view of fin & blade chaeta (specimen no. 11, 10,000 x). Magnifications: A1, A2—7,000 x, A3, E–G—10,000 x, B—6,000 x, C—16,000 x; D1—15,000 x; D2, D3—12,000 x.

FIGURE 28. Serpula hartmanae paratype from the Marshall Islands (U.S. National Museum of Natural History specimen no. USNM 38401). A—Operculum, apical view, 18 blunt radii (slightly damaged), B—Same, lateral view; note constriction with asymmetric boss, C—Tube, showing cross-section; note granular overlay, D—Tube from above, laterally rugose; note three longitudinal ridges, median ridge lower; upper surface rather flat, E1—Collar chaetae, frontal view, appearing somewhat abraded, E2—Collar chaeta, lateral view, F—Abdominal chaetae, G—Abdominal uncinus, F+4. Scales: B–C—1 mm.
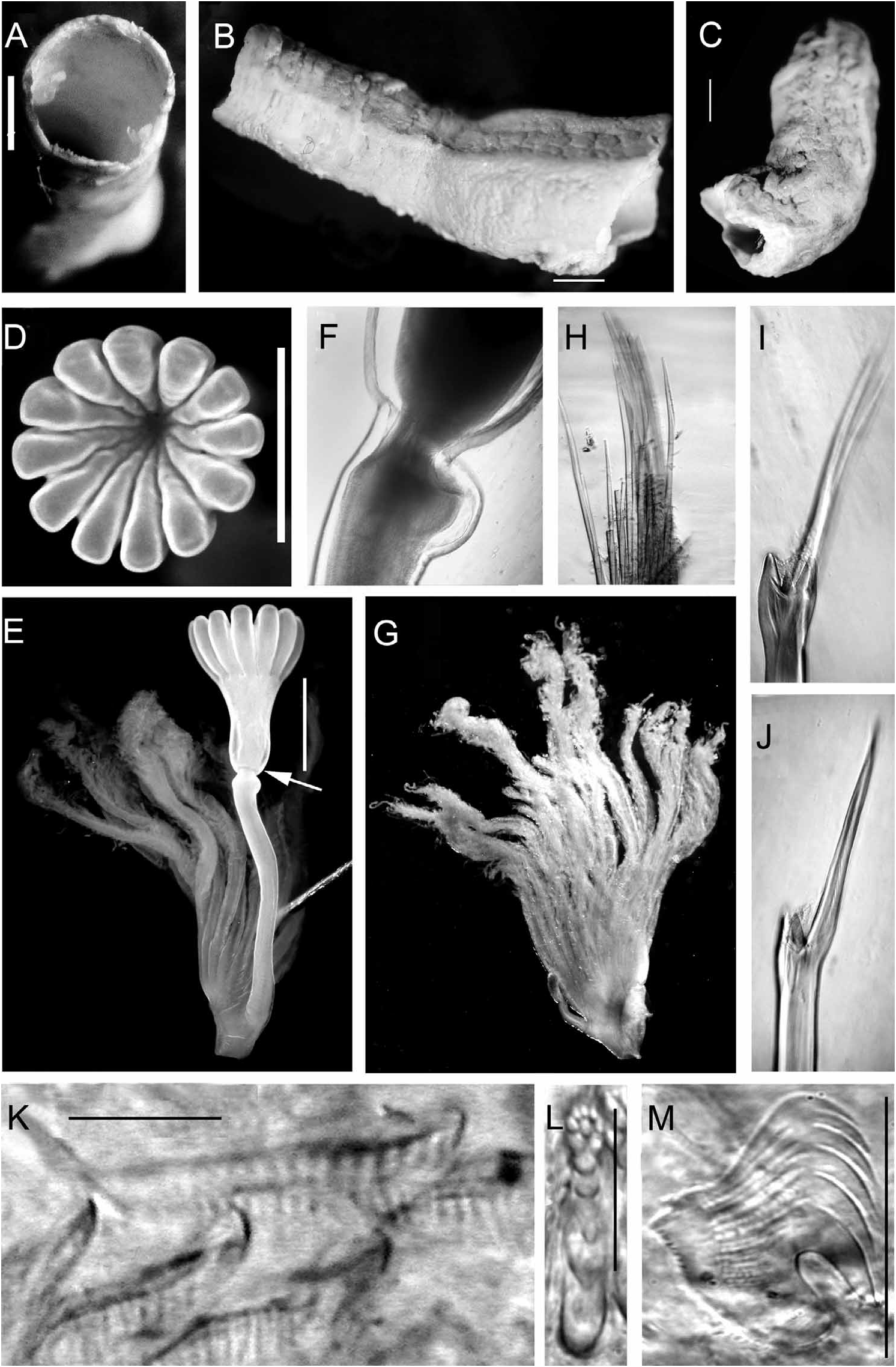
FIGURE 29. Serpula hartmanae from Red Sea, Gulf of Aqaba. Specimen det. as S. concharum in Amoureux et al., 1978: 143, Fig. 11, marked S.c. [TAU-NS-6188], redet. 1990, H.A. ten Hove. A—Cross-section of tube, B—Part of tube, flat surface up, shows 3 longitudinal ridges, middle ridge lower than laterals, C—Part of tube, flat upper part to the right, D—Operculum, apical view, 12 radii, E—Operculum (lateral view) and opercular lobe; arrow designates the peduncular boss proximal to the constriction (also in Fig. 28B), F—View of constriction of opercular peduncle showing characteristic asymmetric proximal boss (enhanced in glycerine mount), G—Branchial lobe, showing pseudoperculum, ca.13 radioles, H—Fascicle of thoracic chaetae (most broken), I, J—Collar chaeta (single unbroken chaeta dissected out from base of fascicle), I—Frontal view, small median tooth between large conical teeth and blade clearly seen, J—Lateral view, K—Abdominal chaetae, L—Posterior? middle abdominal uncinus frontal view (composite picture enables showing rows in focus), detailed dentition, F:1:1:1:1:2:3:3 = F+7/3 rows with maximum number of 3 teeth in the posterior rows, M— Thoracic uncinus, lateral view F+5. Scales: A–E—1 mm, K—10 µm, L, M—50 µm.

FIGURE 30. Serpula hartmanae from the Suez Canal, Great Bitter Lake (Brattström & J.P. Taasen specimen no. 2). A— Tube, showing cross-section; note 3 dorsal longitudinal ridges, tube rather rounded, B—Tube, dorsal aspect, laterally rugose, C—Operculum, apical view, 15 blunt radii, D—Same, lateral view; note constriction with asymmetric boss, E— Whole worm, removed from tube, F—Microscope view of base of operculum with constriction and proximal asymmetric boss (methylene-blue glycerine mount medium), G—Opercular lobe, 11 radioles (in methylene-blue glycerine mount). Scales: A, D—1 mm, E—5mm.

FIGURE 31. Serpula hartmanae from the Suez Canal, Brattström & J.P. Taasen specimen no. 2. A—Collar chaetae, note that two left chaetae have a small accessory tooth between the two large conical teeth and the blade (see also Fig. 32F), B—Thoracic chaetae, C—Abdominal chaetae, D—Thoracic uncini, F+5, E—Abdominal uncini, F+4, F—Radioles with hyaline bodies, G—Enlargement of same. Scales: A, C, G—100 µm, B—1 mm, D, E—10 µm, F—1 mm (estimated from G).

FIGURE 32. Serpula hartmanae from the Suez Canal, Brattström & J.P. Taasen specimen no. 1. A—Tube anterior, B— Tube, 3/4 side-top view, C—Operculum, apical view, 12 radii, D—Operculum, lateral view, E—Branchial radiole showing hyaline bodies, F—Collar chaetae, with 2 conical teeth, with small median tooth visible behind large conical teeth, G—?Abdominal uncinus, lateral view, H—?Abdominal uncinus, ¾ frontal view, I—Abdominal chaetae. Scales: A, B— 1 mm, C, D—0.5 mm, E, G, H—100 µm, F, I—10 µm.
| USNM |
Smithsonian Institution, National Museum of Natural History |
| BM |
Bristol Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Serpula hartmanae Reish, 1968
| Ben-Eliahu, M. Nechama & Ten Hove, Harry A. 2011 |
Serpula hartmanae
| Imajima, M. & Hove, H. A. ten 1986: 2 |
| Imajima, M. & Hove, H. A. ten 1984: 36 |
| Reish, D. J. 1968: 229 |
Serpula sp.
| Hartman, O. 1954: 641 |