
Renicola sp.
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3784.5.5 |
|
publication LSID |
lsid:zoobank.org:pub:95E7BE61-021D-476D-89A9-73F22B39759A |
|
DOI |
https://doi.org/10.5281/zenodo.5062503 |
|
persistent identifier |
https://treatment.plazi.org/id/387F87C7-E414-EE6D-FF34-0CDDAD47A223 |
|
treatment provided by |
Felipe |
|
scientific name |
Renicola sp. |
| status |
|
Renicola sp. “polychaetophila”
( Figs. 1–3 View FIGURES 1–2 View FIGURE 3 )
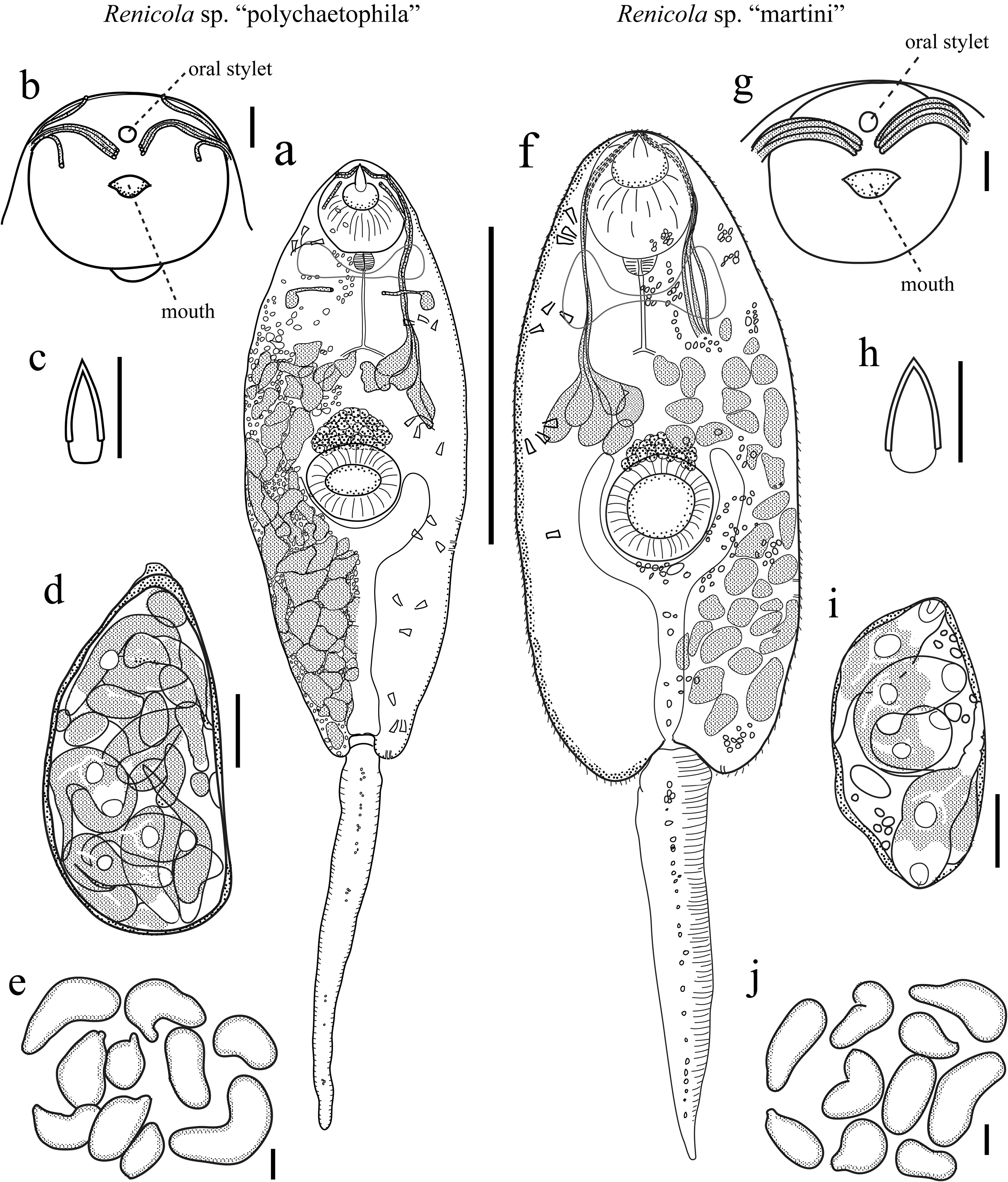
Diagnosis: Distome, leptocercous xiphidiocercariae with penetration gland ducts opening in a 2[(1+3+1)+1] arrangement and no tegmental spines. Cercariae produced by daughter sporocysts comprising a colony that resides in the gonadal and digestive gland regions of the first intermediate host.
Host: Cerithidea californica ( Haldeman, 1840)
Location in host (daughter sporocysts): gonadal and digestive gland regions
Prevalence: 0.7% in 5392 snails dissected in 2012
Locality: Carpinteria Salt Marsh , Santa Barbara County, California, USA (34.4°N, 119.54°W) GoogleMaps
Habitat: Estuaries (intertidal flats, pans, channels)
Dates of collection: May–October 2009
Deposited Material: Cercariae and sporocysts from 5 infections (USNPC nos. 107294–107306).
GenBank accession numbers: CO1 KF512550 View Materials – KF5120552; ITS1 KF512569 View Materials – KF512571 View Materials .
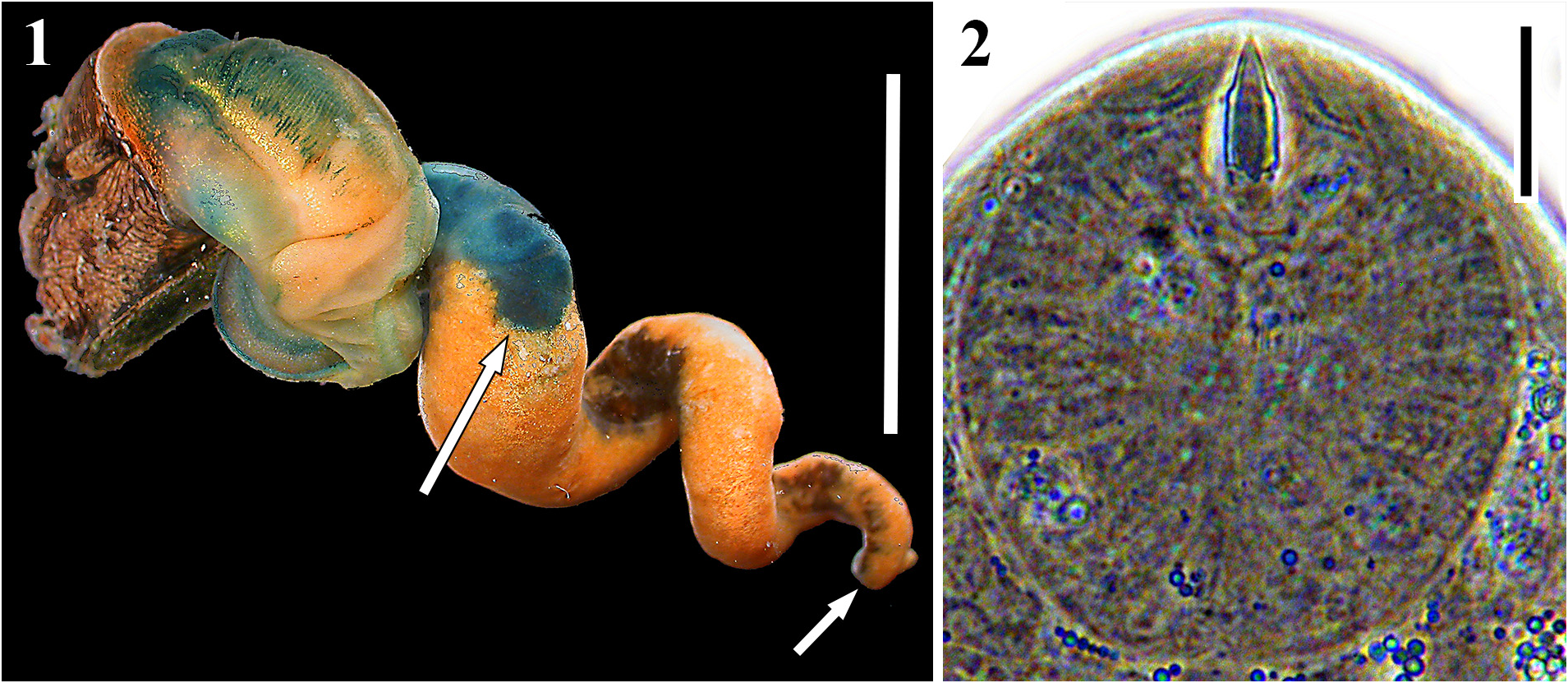
Description: Daughter sporocysts. Body immobile, shape variable, elongate to pyriform to spheroidal, sometimes pinched at one end to form a nipple or neck that appears to mark the birth pore. Tegument translucent, typically covered with thicker paletot that gives sporocysts lemon-cream to orangish appearance and causes them to stick together, and lined on interior by tissue that likely represents germinal cells. Ripe sporocysts with cercariae in various stages of development; germinal balls to developed cercariae distributed in sporocyst with no clear spatial polarity. Developed cercariae impart a whitish coloration to sporocysts under reflected light that mottles with the orange and clear tegument color. Distribution of sporocysts includes gonadal space with some infiltration of digestive gland space. Sporocysts exhibit seasonal variation, wherein sporocysts in developed infections physically regress in winter, becoming thinner-bodied, somewhat flaccid, more orangish, with zero to few developed cercariae; sporocysts ripen up again during spring, becoming more plump, filled with more developed and developing cercariae, and collectively form a more compact mass. Morphometrics and meristics are presented in Table 1 View TABLE 1 and are based on 40 individuals from four infections, while cercaria counts are of the well-developed cercariae from 31 sporocysts from four infections.
Cercariae. Distome, leptocercous xiphidiocercaria. Body elliptic-elongate-ovate in dorsal view, dorso-ventrally flattened, maximal width and dorso-ventral height usually just anterior to ventral sucker; with refringent, varying sized spherules scattered throughout most of interior. Cystogenous glands fill posterior ~¾ of body, densely packed, filled with refringent granules, reflect white light, obscure fine internal anatomy. Oral sucker welldeveloped, round, large, equal-subequal in size with ventral sucker; with small stylet; stylet embedded horizontally at anterior oral sucker, dorsal to mouth, small, usually appears less than ¼ length of oral sucker, sclerotized for 70– 90% of its length, bullet-shaped, tapering anteriorly for about ½ sclerotized length, difficult to carefully observe unless cercaria is flattened. Mouth subterminal; prepharynx absent; pharynx small, ovoid-spheroid, abutting posterior margin of oral sucker; esophagus thin and thin-walled, branching into two ceca approximately midway between oral and ventral suckers near anterior margin of cystogenous gland field; ceca thin and thin-walled, posterior-most extent obscured by cystogenous glands. Ventral sucker positioned at mid-body, well developed, round, protrudes from ventral surface, lacking spines. Tegument thin, apparently with no spines on body or tail (there are potentially extremely minute spines embedded within the tegument); with few, thin sensillae> 10 long observed laterally and dorsally, not counted. Penetration gland bodies extremely difficult to observe given obscuring cystogenous glands, but are primarily clustered ~2/5 into body from anterior edge, anterior extent of cluster close to anterior extent of cystogenous gland field; appear to be five pairs (matching duct number) of unknown number of types. Penetration gland ducts pass anteriorly from glands in bundles of five that pass around lateral margins of oral sucker; each bundle diverges into a 1+3+1 arrangement at lateral and anterolateral edge of oral sucker; one pair exits body slightly dorsal, one pair slightly ventral, and three middle pairs empty just lateroventral to oral stylet tip (the 5 ducts rarely distinguishable from each other before diverge into groups). Additional pair of probable penetration glands present ~1/5 into body from anterior edge, anterior to and more lateral than main penetration gland cluster; each with a single, usually distinct “cross duct” extending medially and ventrally from gland body that opens on ventral body surface. Three pairs of lateral gland ducts of unknown type opening on lateral body margins just posterior to ventral sucker (third frequently not visible); gland bodies not reliably observed. Two pairs of posterior gland ducts of unknown type openings at posterior body margin, one more dorsal to the other (one frequently not readily visible); gland bodies not reliably observed. Excretory bladder prominent, Y-shaped; stem longer than arms; arms embracing posterior half or more of ventral sucker margin; excretory opening at posterior edge of bladder stem. Flame cells in a 2 [(3+3+3) + (3+3+3)] = 36 bilateral arrangement, posterior four groups usually partially obscured by cystogenous glands. Other details of collecting ducts not observed. Cerebral ganglion just posterior of, and extending laterally of, pharynx. Genital primordium a compact mass of cells, adjacent to dorso-anterior margin of ventral sucker, very evident upon acetocarmine staining. Tail attached terminally at posterior body margin in socket, simple, unadorned, slender, cylindrical in cross-section, gradually tapering, can extend and contract, approximately equal in length to body when extended, with finely annulated tegument overlying muscle layers, with scattered, refringent spherules along mid-line. Morphometrics are presented in Table 1 View TABLE 1 ; most are based on 40 individuals from four infections, while stylet measurements come from 14-18 individuals from three to four infections.
Behavior: Cercariae swim by widening and flattening the mid- and posterior-body, flexing posterior body ventrally to partly overlap ventral sucker, often also slightly flexing or protruding anterior body, and lashing the tail left to right to move in a general anterior direction. Cercariae do not clearly respond to light.
1 Observations on additional, fresh material indicates that sporocysts can be larger than indicated by material used for morphometrics.
2 Counts of well-developed cercariae.
3 Stylet length-t is the total length of the stylet (sclerotized and non-sclerotized portions)
4 Stylet length-s is the length of the sclerotized portion of the stylet
Other biology: The trematode appears to usually infect the host by crossing the mid-gut, as expected given its taxonomic affiliation, and as evidenced by residual sporocyst or germinal material in the parenchymous tissue surrounding the mid-gut.
Developed infections parasitically castrate the host and usually appear to almost completely replace snail gonadal tissues. However, the seasonal (winter) regression, where cercaria production is greatly decreased and sporocysts become somewhat flaccid, is associated with infiltration of the parthenita colony by snail digestive gland tissue and sometimes a small amount of snail gonadal tissue regeneration (but there is no sign that snail reproduction takes place). These observations are consistent with the information provided in Hechinger et al. (2009) that the aggregate sporocyst mass takes about 20% of the infected host soft tissue mass in summer and 16% in winter. That study may have pooled this species and Renicola sp. “martini”, but the two species appear to be very similar concerning the way they grossly use host space.
Infections are sometimes encountered that are in the process of being invaded by other trematode species, particularly those with rediae, which can be observed ingesting the sporocysts and cercariae of Renicola sp. “polychaetophila”. This is consistent with the relatively low ranking of this species in the interspecific dominance hierarchy characterizing the guild of trematodes infecting the California horn snail ( Kuris 1990; Sousa 1993).
Field surveys and preliminary laboratory exposure experiments indicate that the cercariae encyst as metacercariae in the hemocoel of various estuarine polychaetes (Hechinger and Smith, unpublished data; see list of probable hosts in Hechinger et al. (2011a)).
The final host species are not determined, but almost certainly include birds that prey on polychaetes.
Geographic distribution: This species may occur throughout the entire range of the California horn snail, from North of San Francisco Bay (California, USA) to northern Peru ( Keen 1971; Miura et al. 2010). However, we have only positively identified it from snails from southern San Francisco Bay to the Estero de Punta Banda (northern Baja California, Mexico). In addition, preliminary genetic data indicates that this species occurs in the Atlantic Cerithidea pliculosa (Menke) (sister species to the California horn snail), in the Gulf of Mexico and the Yucatan Peninsula (Miura et al. unpublished data).
Etymology: The potential specific epithet, “polychaetophila” (“polychaete loving”), reflects the second intermediate host use of this species.
Remarks: Renicola sp. “polychaetophila” fits well within the emended diagnosis for renicolid cercariae (above), and the molecular results (below) further indicate that it is embedded within Renicola .
Table 2 lists attributes of previously described renicolid xiphidiocercariae. The arrangement of the penetration gland-bodies and ducts readily distinguishes Renicola sp. “polychaetophila” from most other renicolid xiphidiocercariae. With one possible exception (below), it is the only species that has a 2[(1+3+1)+1] arrangement of penetration gland-duct openings, where the gland bodies for the five anterior-most duct openings are clustered together and lie within the cystogenous-gland field, while the gland bodies for the posterior-ventral “cross-duct” openings are solitary and anterior to the penetration-gland field. Of the four species that are at least roughly similar to Renicola sp. “polychaetophila” in having penetration gland ducts that diverge to open away from the stylet, Cercaria caribbea XXXIII, Cable, 1956 is most similar. However, C. caribbea XXXIII does not appear to have the pair of “cross-duct” penetration gland bodies isolated anteriorly from the main cluster of penetration gland bodies (which lie in the obscuring cystogenous gland field) and it also has tegmental spines whereas Renicola sp. “polychaetophila” does not (or at least it does not have any readily apparent tegumental spines).
Specimens of Renicola sp. “polychaetophila” appear to have been included in the material that Hunter used to describe “ Cercaria cerithidia 19” in her unpublished 1942 dissertation. However, it seems her material also included Renicola cerithidicola Martin, 1971 (a non-styleted species with sporocysts in the mantle area), perhaps as mixed-species infections with Renicola sp. “polychaetophila”, as her description fuses characteristics of both those species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |