
Athanopsis brevirostris Banner & Banner, 1981
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5383.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:7E434B40-6346-4E6D-BC55-38EBAE24BD52 |
|
DOI |
https://doi.org/10.5281/zenodo.10392021 |
|
persistent identifier |
https://treatment.plazi.org/id/386ABE42-FFA0-FFBC-FF06-AD0B2F63F9AC |
|
treatment provided by |
Plazi |
|
scientific name |
Athanopsis brevirostris Banner & Banner, 1981 |
| status |
|
Athanopsis brevirostris Banner & Banner, 1981 View in CoL
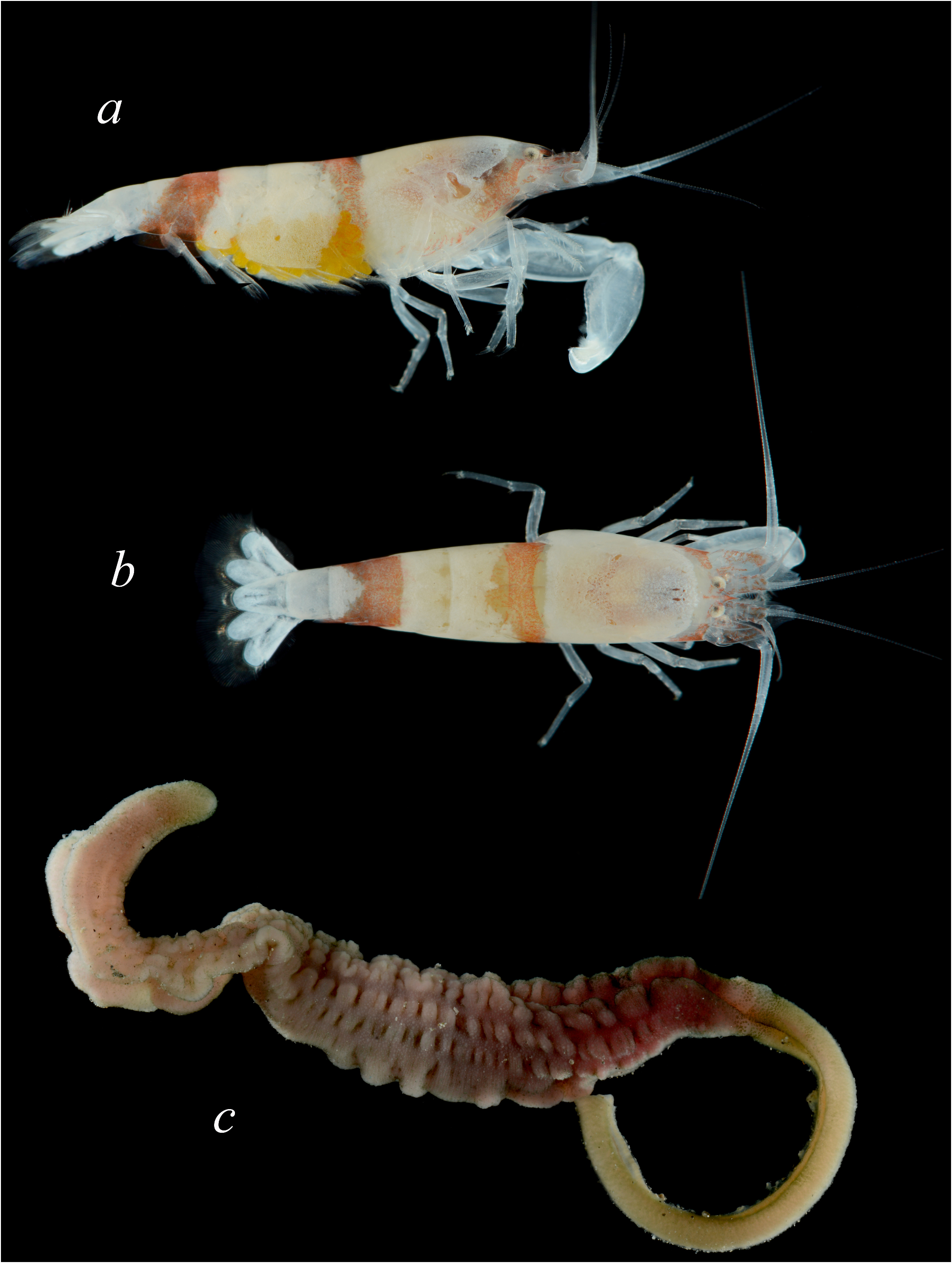
( Fig. 8a, b View FIGURE 8 )
Athanopsis brevirostris Banner & Banner 1981: 45 View in CoL , fig. 5; Anker 2012: 51 View Cited Treatment , figs. 2–4, 7A, B.
Material examined. 1 ovig. female (cl 5.0 mm), FLMNH UF 71444, Saudi Arabia, Thuwal, near KAUST King Abdullah Monument, 22°20’26.2”N / 39°05’15.1”E, shallow subtidal sandflat with seagrass and rubble patches, depth 0.5–1.5 m, suction pump, in echiuran burrow, leg. A. Anker, 20.01.2023 [fcn AA-22-497]; 1 male (cl 4.1 mm), 1 ovig. female (cl 4.4 mm), FLMNH UF 71426 , same collection data as for previous specimen [fcn AA-22-474] ; 1 male (cl 3.6 mm), FLMNH IUF 71429 , same collection data as for previous specimens [fcn AA-22-489] ; 1 male (cl 3.9 mm), FLMNH UF 71441 , same collection data as for previous specimens [fcn AA-22-467]; 1 ovig. female (cl 4.4 mm), FLMNH UF 71442 , same collection locality as for previous specimens, leg. A. Anker, 21.01.2023 [fcn AA-22-500] ; 1 male (cl 4.4 mm), FLMNH UF 71427 , same collection data as for previous specimen [fcn AA-22-511] ; 1 female (cl ~ 3.5 mm), FLMNH UF 36637 , Saudi Arabia, Farasan Islands , Marca Island, 18°13’14.0”N / 41°19’27.8”E, subtidal sandflat, depth 5–10 m, suction pump, in burrow of unknown host, leg. A. Anker et al., 06.03.2013 [fcn BDJRS-1756] GoogleMaps .
Comparative material. Athanopsis saurus Anker, 2011: 1 female (cl 3.6 mm), MNHN-2018-571, New Caledonia, Koumac GoogleMaps , Pointe Pandop, Expedition Koumac 2.1 sta. KM601 , 20°35’S / 164°16.5’E, partly exposed sand-mud intertidal with coral rubble fragments, depth 0–0.2 m, suction pump, in echiuran burrow, leg. A. Anker, 08.09.2018; 1 female (cl 5.0 mm), MNHN-2018-1012, New Caledonia, Saint-Mathieu , Expedition Koumac 2.1 Miscellanea sta. KM606 , 20°22.8’S / 164°35’E, shallow sandflat with sparse seagrass near Pouébo river estuary and mangroves, depth 0–0.5 m, suction pump, in echiuran burrow, leg. A. Anker & Z. Ďuriš, 18.09.2018 GoogleMaps ; 1 ovig. female (cl 4.9 mm), MNHN-2018-978, same collection data as for previous specimen GoogleMaps ; 1 male (cl 4.3 mm), MNHN- IU-2018-974, same collection data as for previous specimens GoogleMaps .
Description. See Banner & Banner (1981) for original description and illustrations, and Anker (2012) for additional illustrations of the Madagascan material; colour pattern of the Saudi Arabian specimens is shown in Fig. 8a, b View FIGURE 8 .
Colour in life. Described in detail in Anker (2012: 53, fig. 7A, B); see also Fig. 8a, b View FIGURE 8 .
Distribution. Red Sea: Eritrea: Dahlak Archipelago ( Banner & Banner 1981), Saudi Arabia: Thuwal, Farasan Islands (present study); Madagascar: Nosy-Bé; Japan: Okinawa ( Anker 2012).
Ecology. Most Saudi Arabian specimens of A. brevirostris were collected together with echiuran hosts, a large reddish species possibly belonging to either the genus Ochetostoma Rüppel & Leuckart or the related genus Listriolobus Fischer ( Thalassematidae ) ( Fig. 8c View FIGURE 8 ). Some of the previously reported specimens of A. brevirostris were also extracted from burrows of Ochetostoma sp. or unknown hosts, however, the holotype came from a poisoning station ( Banner & Banner 1981; Anker 2012). Thus, the present evidence suggests that all species of Athanopsis live in symbiosis with echiurans ( Miya 1980; Berggren 1991; Anker 2011, 2012; Marin et al. 2014; present study).
Remarks. Anker (2012) separated A. brevirostris from the closely related A. saurus (type locality: Lizard Island, Great Barrier Reef) by the following four features: (1) rostrum short, with lateral margins broadly concave in A. brevirostris vs. longer, with lateral margins proximally convex in A. saurus ; (2) second antennular peduncle much wider than long in A. brevirostris vs. only slightly wider than long (almost square-shaped) in A. saurus ; (3) major chela palm with small field of setae distally in A. brevirostris vs. with more extensive setation in A. saurus ; and (4) colour pattern with more white on the second and third pleonites and less red on the carapace in A. saurus . As pointed out by Anker (2012), in the holotype of A. brevirostris , as well as in the Madagascan material assigned to that species, the merus of the third to fifth pereiopods has a small distolateral tooth, which was not overlooked by Banner & Banner (1981); this tooth is present, and sometimes quite conspicuous, in all Saudi Arabian specimens.
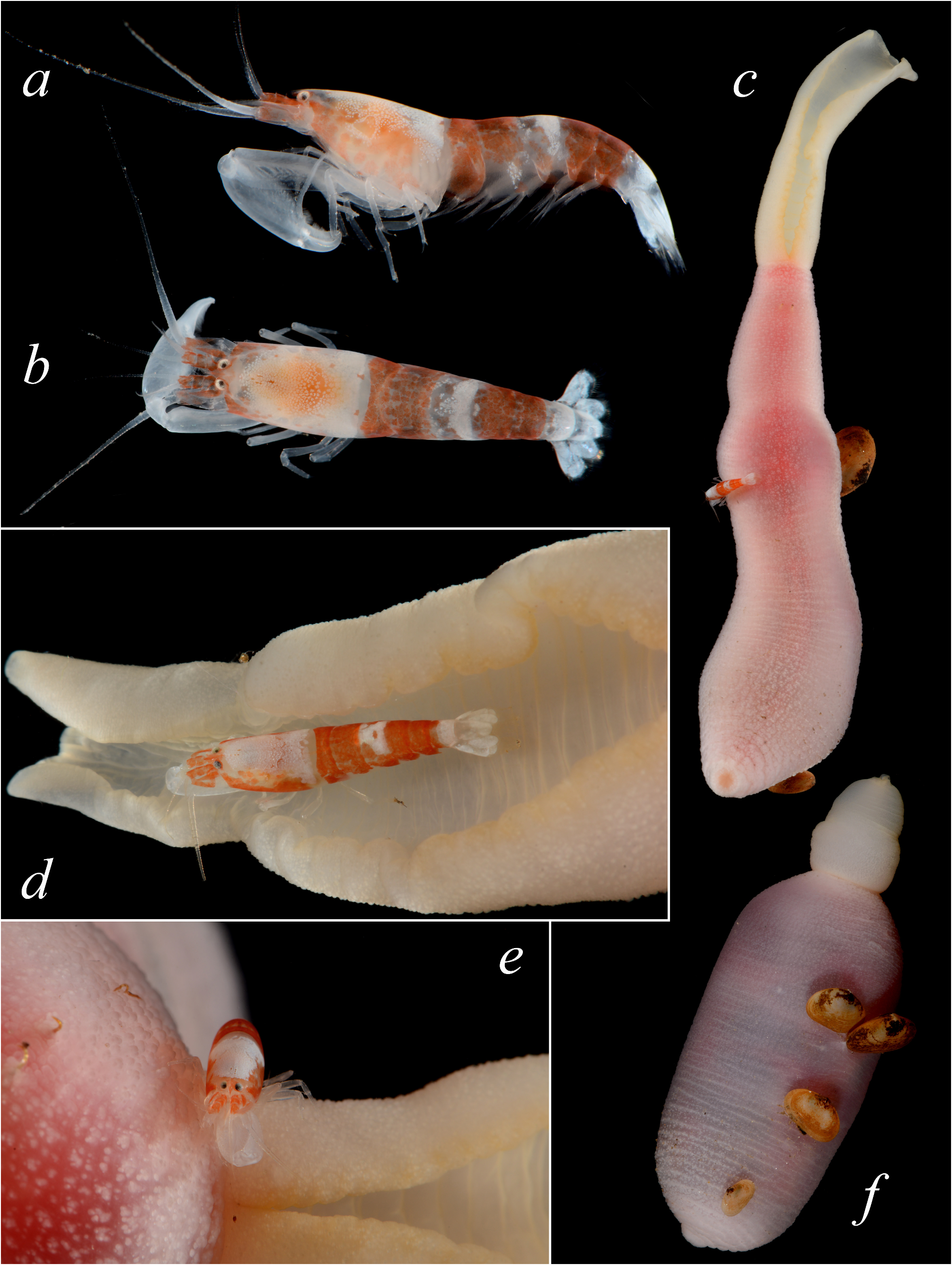
The New Caledonian material is morphologically somewhat intermediate between A. brevirostris and A. saurus , especially in the features of the rostrum and major chela, raising at least the possibility that the two species may be synonymous. Indeed, the aforementioned morphological differences between A. brevirostris and A. saurus are slight and somewhat subjective, and need to be verified when more material becomes available. In addition, the colour pattern of the Saudi Arabian specimens of A. brevirostris ( Fig. 8a, b View FIGURE 8 ) appears to be closer to the pattern of A. saurus from Lizard Island (Anker 2011: fig. 8G, H; Anker 2012: fig. 7E), whereas the colour pattern of the Madagascan specimens reported as A. brevirostris ( Anker 2012: fig. 7A, B) is very similar to that of the New Caledonian specimens herein identified as A. saurus ( Fig. 9a–e View FIGURE 9 ). Thus, the present material largely eliminates the colour differences between the two species listed by Anker (2012). Nonetheless, while contrasting published colour photographs of A. brevirostris and A. saurus (Anker 2011, 2012) with those available for the present material ( Figs. 8 View FIGURE 8 , 9 View FIGURE 9 ), the author noticed one colour difference that could reveal as diagnostic. In the shrimps identified as A. brevirostris from the Red Sea (Farasan Islands, Thuwal), Madagascar (Nosy Bé), and Japan (Okinawa), the carapace is typically white or white locally speckled with small red chromatophores, especially on the anterolateral surface ( Fig. 8a View FIGURE 8 ; Anker 2012: fig. 7A, B; Goto & Kato 2012: fig. 1h). In the holotype of A. saurus from Australia (Lizard Island) and specimens from New Caledonia (Koumac, Pouébo) herein identified as A. saurus , the carapace is white with large patches of red chromatophores on the anterolateral surface ( Fig. 9a, d View FIGURE 9 ; Anker 2011: fig. 8G; Anker 2012: fig. 7E). It also appears that the red patches on the pleon of A. saurus extend to additional pleonites. Importantly, A. brevirostris and A. saurus were recovered as sister species in the molecular phylogeny of Chow et al. (2021: fig. 3), where the former was represented by a specimen from Okinawa (listed in Anker 2012), and the latter by a specimen from New Caledonia (MNHN-IU-2018-978). The genetic distance between them was not significant, but sufficient to be considered as interspecific (3.1% for 16S; genetic distance presently unavailable for COI). The next step would be to compare sequences from the holotype of A. saurus and the western Indian Ocean (Red Sea, Madagascar) material of A. brevirostris , to draw the final conclusion about the taxonomic status of the former species.
| FLMNH |
Florida Museum of Natural History |
| UF |
Florida Museum of Natural History- Zoology, Paleontology and Paleobotany |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Caridea |
|
Family |
|
|
Genus |
Athanopsis brevirostris Banner & Banner, 1981
| Anker, Arthur 2023 |