
Dohrnemesa kuarajucassaba, Gil-Santana & Ferreira, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4338.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:6043549C-C784-4EAF-9BC6-E009027B4F47 |
|
DOI |
https://doi.org/10.5281/zenodo.6008792 |
|
persistent identifier |
https://treatment.plazi.org/id/37738795-C716-FFDC-45F4-F9DFFC48FAF4 |
|
treatment provided by |
Plazi |
|
scientific name |
Dohrnemesa kuarajucassaba |
| status |
sp. nov. |
Dohrnemesa kuarajucassaba View in CoL sp. nov.
( Figs. 1–79 View FIGURES 1 – 3 View FIGURES 4 – 8 View FIGURES 9 – 18 View FIGURES 19 – 23 View FIGURES 24 – 26 View FIGURES 27 – 29 View FIGURES 30 – 34 View FIGURES 35 – 40 View FIGURES 41 – 48 View FIGURES 49 – 51 View FIGURES 52 – 54 View FIGURES 55 – 59 View FIGURES 60 – 63 View FIGURES 67 – 72 View FIGURES 73 – 79 , 82–83 View FIGURES 80 – 83 )
Diagnosis. Somewhat larger: while length of other Dohrnemesa species ranges from 10 to 16 mm, the total length of D. kuarajucassaba sp. nov. is 18.5–18.8 mm (males) to 21.1–23.0 mm (females). The males most closely resemble the male of D. lanei . The two species can be readily separated by the following characteristics: 1— postocular portion of head with a median distinct tubercle ( D. lanei ) or an oval small shallow tumescence ( D. kuarajucassaba sp. nov.); 2—Pcu meeting discal cell somewhat distad ( D. kuarajucassaba sp. nov., Figs. 27–29 View FIGURES 27 – 29 , 31 View FIGURES 30 – 34 ) or proximad ( D. lanei ) of the apex of basal cell on forewing; 3—sternites with several whitish markings in D. kuarajucassaba sp. nov. ( Fig. 36 View FIGURES 35 – 40 ) and uniformly blackish in D. lanei . The females of D. kuarajucassaba sp. nov. differ from all known females of Dohrnemesa by their forewings greatly reduced, reaching about middle of abdomen ( Figs. 49–51 View FIGURES 49 – 51 , 64, 74), extremely reduced hind wings (Fig. 66), and abdominal segments IV–VI greatly widened ( Figs. 73–76 View FIGURES 73 – 79 ).
Description. Male. Figures 1–48 View FIGURES 1 – 3 View FIGURES 4 – 8 View FIGURES 9 – 18 View FIGURES 19 – 23 View FIGURES 24 – 26 View FIGURES 27 – 29 View FIGURES 30 – 34 View FIGURES 35 – 40 View FIGURES 41 – 48 . Measurements: Table 1.
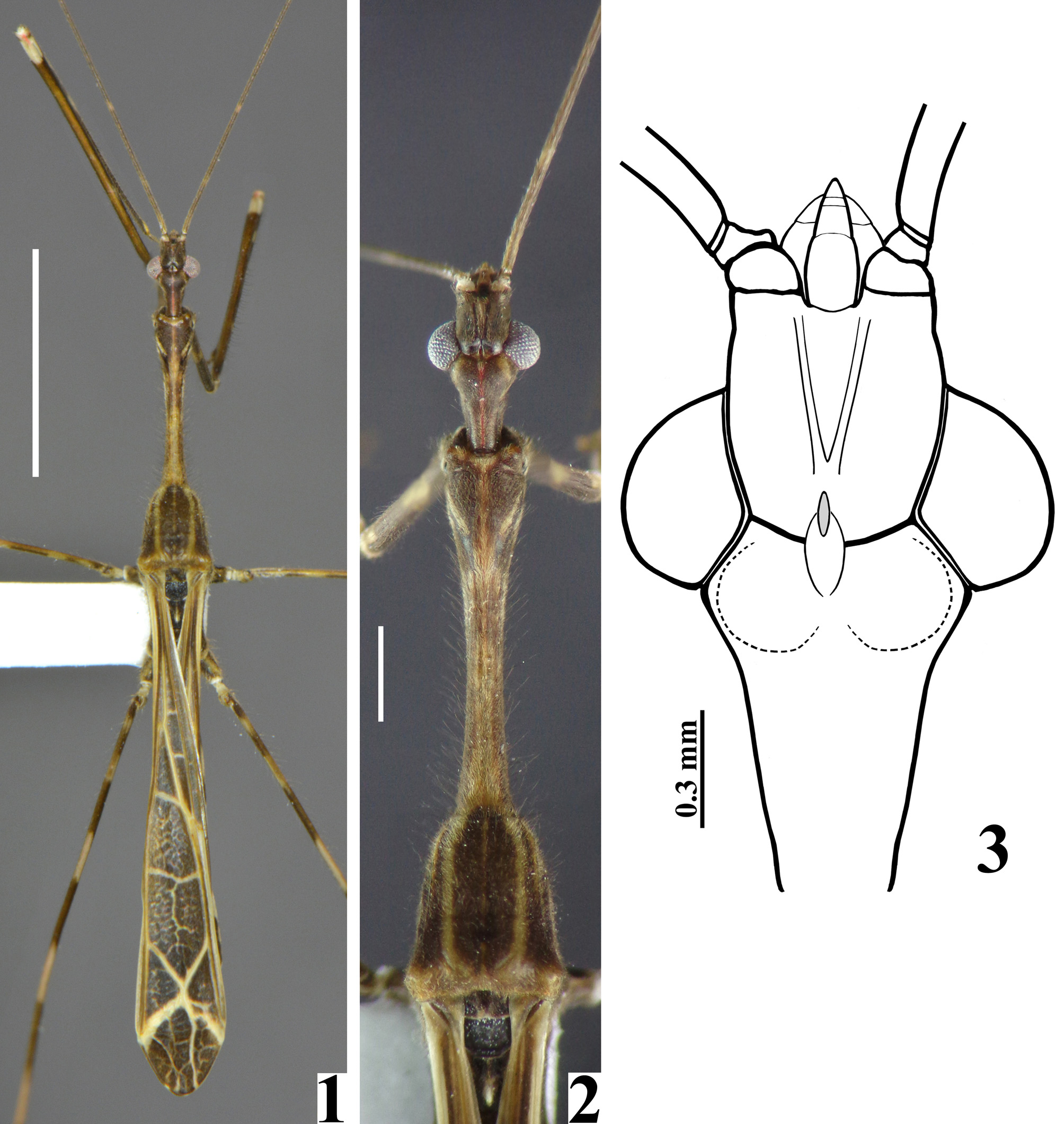
Coloration: black to brownish black, with pale, yellowish or whitish markings or portions ( Figs. 1 View FIGURES 1 – 3 , 4 View FIGURES 4 – 8 ). Head black to brownish black; dorsally somewhat pale at midline on anterior half of anteocular region, but blackish on area just behind clypeus; on postocular region, a midlongitudinal thin reddish stripe; clypeus, labrum, apices of antenniferous tubercles, apices of labial segments II and III and base of segment IV pale to pale white; laterally, pale yellowish spots: a somewhat rounded spot adjacent to upper third of eye, anteriorly, and a pair of somewhat more elongated spots going from posterior rim of eye, at its upper half, to approximately midportion of postocular region ( Figs. 2 View FIGURES 1 – 3 , 5 View FIGURES 4 – 8 ). Antenna brownish to black-brownish with all intersegmental joints paler and narrow pale annuli, as follows: segment I with five annuli and apical extremity pale, the first (subbasal) annulus always longer, approximately five times longer than the width of the segment, the next three annuli about twice to four times as long as the width of the segment; and the fifth (subapical) annulus sometimes shorter, only somewhat longer than the width of the segment ( Fig. 6 View FIGURES 4 – 8 ). Segment II with three annuli, two at basal half and one somewhat distal to midpoint, the first about three to four times, second five to seven times and third three to seven times longer than the width of the segment (the variation in length of these annuli was even observed between the two antenna of the same specimen) ( Fig. 7A–B View FIGURES 4 – 8 ); apical extremity pale ( Fig. 8 View FIGURES 4 – 8 ). Segment III with apical extremity pale ( Fig. 8 View FIGURES 4 – 8 ). Segment IV mostly dark to pale yellowish with the subbasal and subapical portions dark brownish; basal extremity sometimes even paler ( Fig. 8 View FIGURES 4 – 8 ). Thorax: dark brownish to brownish black ( Figs. 1–2 View FIGURES 1 – 3 , 4–5 View FIGURES 4 – 8 ). Anterior collar at midportion, margin of prothoracic supracoxal lobe and small area above it pale. Pronotum with a dorsal median longitudinal pale line; thinner on fore lobe, wider on anterior fifth and posterior half of petiole and much thinner on hind lobe, not reaching the posterior margin of the latter ( Figs. 1–2 View FIGURES 1 – 3 ). Additionally, on first half, this pale line has a reddish tinge and is somewhat enlarged towards posterior fifth of petiole ( Fig. 2 View FIGURES 1 – 3 ). Hind lobe with a pair of pale submedian, subparallel longitudinal stripes that do not reach posterior margin, ending approximately at the level of anterior half of humeral tubercles; laterally they are accompanied by another pair of thinner shorter pale stripes, which end before the level of the humeral tubercles ( Figs. 1–2 View FIGURES 1 – 3 ). On fore lobe, a curved thin pale stripe running from lateral margin anteriorly and ending near, but not meeting the midlongitudinal pale stripe dorsally; below this stripe another shorter stripe that does not reach the level of the anterior and posterior extremities of the other stripe ( Figs. 1–2 View FIGURES 1 – 3 , 5 View FIGURES 4 – 8 ). At basal third of petiole, laterally, linear ill-defined pale markings. Posterior margin of hind lobe pale ( Figs. 1–2 View FIGURES 1 – 3 ). Lower margin of hind lobe of pronotum pale, forming a curved stripe, somewhat larger at basal half, ending at level of humeral tubercles. Humeral tubercles somewhat pale on central portion, dorsally ( Figs. 1– 2 View FIGURES 1 – 3 ). Spine of scutellum dark with apex pale; spine of metanotum pale at approximately distal half ( Figs. 4 View FIGURES 4 – 8 , 19–20 View FIGURES 19 – 23 ). Meso and hind supracoxal lobes with margins pale. Mesopleura with two and metapleura with one whitish spots on posterior portion. Legs: black to brownish black, with the following yellowish pale to whitish markings or portions: fore coxa with a large submedian proximal annulus and a pair of subapical small pale spots on inner and outer surfaces; apical margin pale ( Fig. 21 View FIGURES 19 – 23 ). Mid and hind coxae with a pair of pale spots on upper surface and apical extremities pale too. Fore trochanter with a pale spot on middle third laterally. Mid and hind trochanters pale on approximately basal third. Fore femora variably paler on dorsal surface, with a pair of pale spots on inner and outer surfaces of basal half, and an apical larger whitish spot, which almost covers all the segment but is absent on ventral surface and therefore does not form a complete ring ( Fig. 21 View FIGURES 19 – 23 ); anteroventral series with some and posteroventral series with most of the large and medium-sized processes with their basal portions and the area surrounding their insertion pale to whitish. Mid and hind femora with basal extremity pale; a subbasal narrow pale spot that does not form a complete ring; five large pale annuli along the segment, the more proximal of them generally narrower than the others, while all are generally narrower than the subsequent dark area; apices whitish for a distance somewhat longer than the preceding pale annulus ( Fig. 24 View FIGURES 24 – 26 ). Basal extremity of tibiae generally with reddish tinge, dorsally. Fore tibia with a pair of pale whitish spots on inner and outer surfaces subbasally, followed by two whitish annuli on basal half and a pair of very small pale yellowish spots on inner and outer surface, somewhat distal from midpoint ( Fig. 21 View FIGURES 19 – 23 ). Mid and hind tibiae mostly pale with three large darkened annuli on approximately basal half; apical extremities darkened too ( Figs. 25–26 View FIGURES 24 – 26 ). Additionally, mid tibiae with a small subbasal dorsal darkened spot. Tarsi darkened. Forewing mostly black to brownish black with basal third of area over costal vein largely pale to whitish; most veins and area surrounding them are, to a variable extent, pale or whitish; inner margin, from the level of Pcu to a point somewhat distally to the level of the apical inner angle of the discal cell, whitish. Discal cell with the following pale markings: diffuse texture formed by small irregular pale whitish spots and short lines; a transverse somewhat curved stripe crossing the cell at middle third and an additional interrupted or rippled transverse stripe at its distal third in the paratypes; a tortuous thin longitudinal approximately median line that runs along the cell to a variable extent. Two larger oblique whitish stripes near tip of forewing, which because of their continuation with veins enclosing apex of discal cell, a whitish figure somewhat similar to an “X” at the distal third is formed; below this “X”, a similar diffuse texture as described above and a longitudinal submedian whitish line, that runs from this “X” to the apical margin ( Figs. 1 View FIGURES 1 – 3 , 27–28 View FIGURES 27 – 29 ). Hind wings hyaline, somewhat more pigmented on distal third above R+M vein, on areas adjoining distal veins and on anal lobe ( Figs. 33– 34 View FIGURES 30 – 34 ). Abdomen brownish to black brownish ( Figs. 35–36 View FIGURES 35 – 40 ); tergites IV–VI with an inconspicuous scattered irregular pattern of pale and grayish markings; tergite VII with posterior margin pale yellowish; connexivum with basal, submedian and apical whitish spots on segments III–VI, the basal spots moderately larger; on segment VII a subbasal whitish spot ( Fig. 35 View FIGURES 35 – 40 ); spiracles and the area surrounding them whitish; sternites with the following whitish markings: a pair of sublateral thin stripes on the distal portion of sternite III; distal half of sternite IV with a sublateral pair of small spots at midportion and a pair of submedian tortuous stripes at distal third; sternites V–VI with submedian tortuous stripes, one pair of which on approximately basal third and two pairs of them on distal third of the segments; a pair of lateral distal spots on sternite VII; a pair of pale rounded apicolateral spots on sternite VIII ( Fig. 36 View FIGURES 35 – 40 ). External male genitalia: pygophore blackish and pale on exposed and hidden portions, respectively; parameres pale, with their apices somewhat darkened.
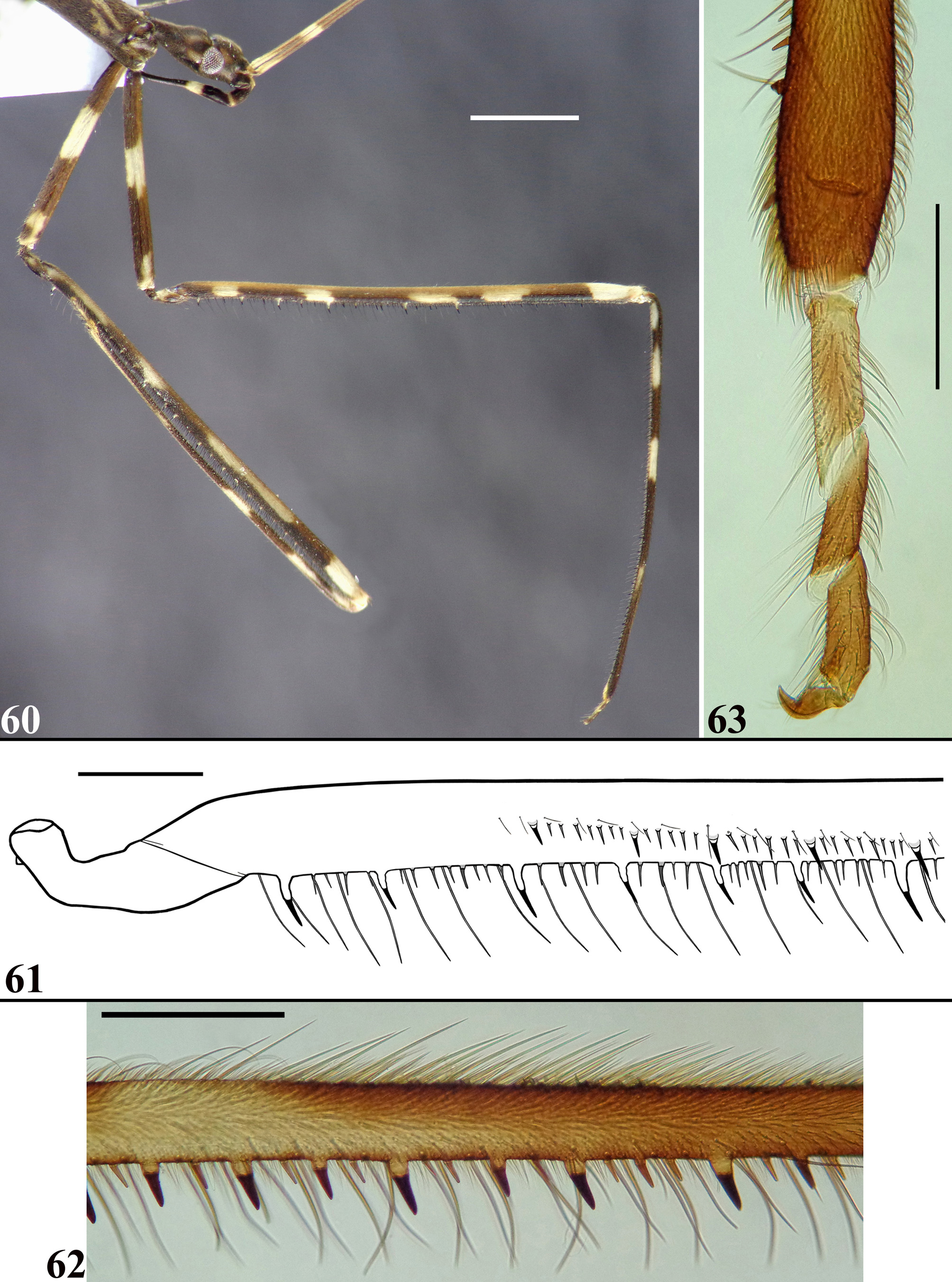
Vestiture: body covered with a very dense pubescence formed by thin, short, adpressed setae and variably with scattered longer setae; generally, they are paler or darker, similar to the coloration of the integument in which they are inserted. In some exceptions mentioned below, groupings of whitish adpressed setae form whitish markings on areas with brownish integument. Head: mostly covered by a pubescence formed by adpressed short thin setae, with scattered longer setae on dorsal region and gula. Glabrous areas on head: transverse sulcus, a declivous area immediately anterior to it and a pair of submedian slightly divergent thin stripes running from midportion of transverse sulcus to a point near medial margin of antenniferous tubercles. Antenna: segments I–II densely covered with pubescence formed by short thin curved setae ( Figs. 9–14 View FIGURES 9 – 18 ); segment I with a few longer thin scattered somewhat curved setae on outer surface ( Figs. 10–11 View FIGURES 9 – 18 ); a few (eleven to fifteen) very much thinner longer isolated elements (interpreted as trichobothria) on segment II, seven to ten of them in its basal portion, implanted before the subbasal proximal pale annulus; others in more distally positions; segments III–IV covered with a pubescence formed by somewhat less numerous, short, straight, obliquely inclined or somewhat curved, thin setae ( Figs. 15–18 View FIGURES 9 – 18 ). Labium: segment II (first visible) with scattered moderately short adpressed setae; segment III and IV almost glabrous with scattered straight erect and some adpressed short setae, somewhat more numerous on dorsal region. Thorax: besides the short dense pubescence and long thin setae, there are several even longer, somewhat thicker, erect setae on pronotum, meso and metasternum ( Figs. 2 View FIGURES 1 – 3 , 5 View FIGURES 4 – 8 ). A glabrous shining curved stripe running between the curved pale stripes of fore lobe of pronotum ( Fig. 5 View FIGURES 4 – 8 ). Meso and metapleura with a lateral somewhat large stripe formed by whitish pubescence. Spines of scutellum and metanotum with numerous moderately short, thin setae. Forewing with a few scattered short thin setae at basal portion and a few scattered somewhat longer ones along costal vein. Hind wing glabrous. Legs generally covered by dense short adpressed pubescence and several long thin setae; the setae generally dark, somewhat paler at apices of mid and hind femora. Mid and hind coxae with three glabrous linear areas on basal two-thirds ventrally, and a small basolateral glabrous area too. Fore femur: posteroventral series beginning at the base of the article, composed of about twelve to fifteen large and medium-sized spiniferous processes, the most basal of which with its apex inclined toward apex of article; among these large processes, approximately seventy to seventy-eight small spiniferous processes; towards apical portion of fore femur, the large and then the small processes become somewhat smaller and absent at approximately distal fourth. Length of larger processes combined with apical spines about three-fourths of diameter of segment. A sparse series of very long and strong setae accompanies the posteroventral series; these setae reach the apex of the femur, where they are somewhat shorter. Anteroventral series beginning somewhat apically to posteroventral series, not interrupted at base, composed of fifteen to twenty-two spiniferous tubercles; among these large processes, approximately 103 to 105 sharp small spines, with an additional nine to ten small spines at the apex of the series. A sparse series of short, thin setae accompanies the anteroventral series; these setae reach the apex of the femora; the setae are even somewhat shorter at basal portion. Fore tibiae with numerous stiff setae on subapical dorsal depression ( Fig. 22 View FIGURES 19 – 23 ); ventral surface with numerous small spines, among which, with exception of the basal fifth of the article, about 29 to 31nine larger spiniferous processes, subequal in size, most of which with lighter base and darkened spiny distal portion; at both sides of ventral surface, beside these processes, series of long and strong setae ( Fig. 22 View FIGURES 19 – 23 ); on apical extremity, a cluster of stiff somewhat curved golden setae, ventrally ( Fig. 23 View FIGURES 19 – 23 ). Tarsi covered with moderately long setae ( Fig. 23 View FIGURES 19 – 23 ). Abdomen: covered with dense adpressed pubescence and scattered thin long setae, which become more numerous on last two or three segments and genital segments.
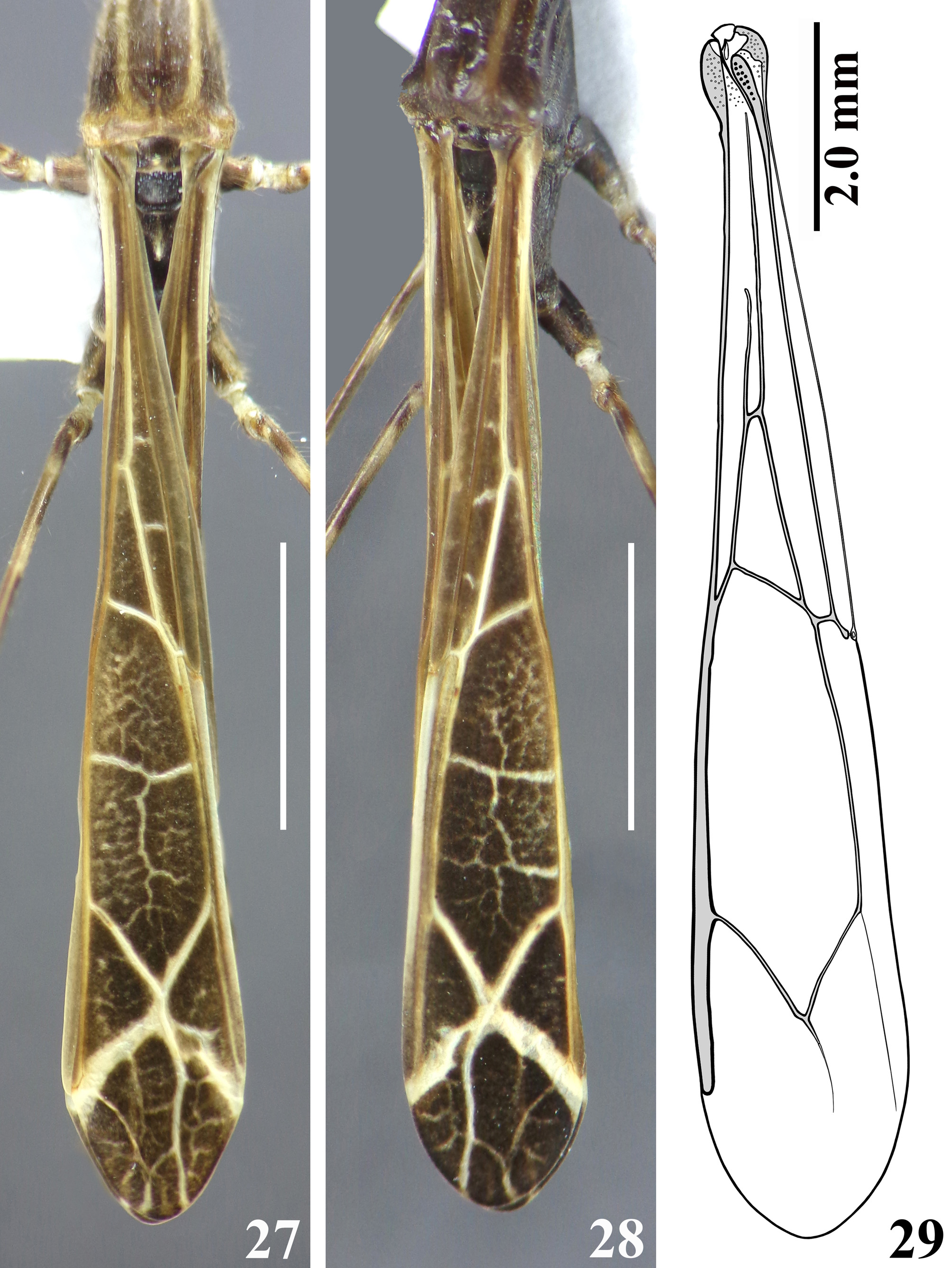
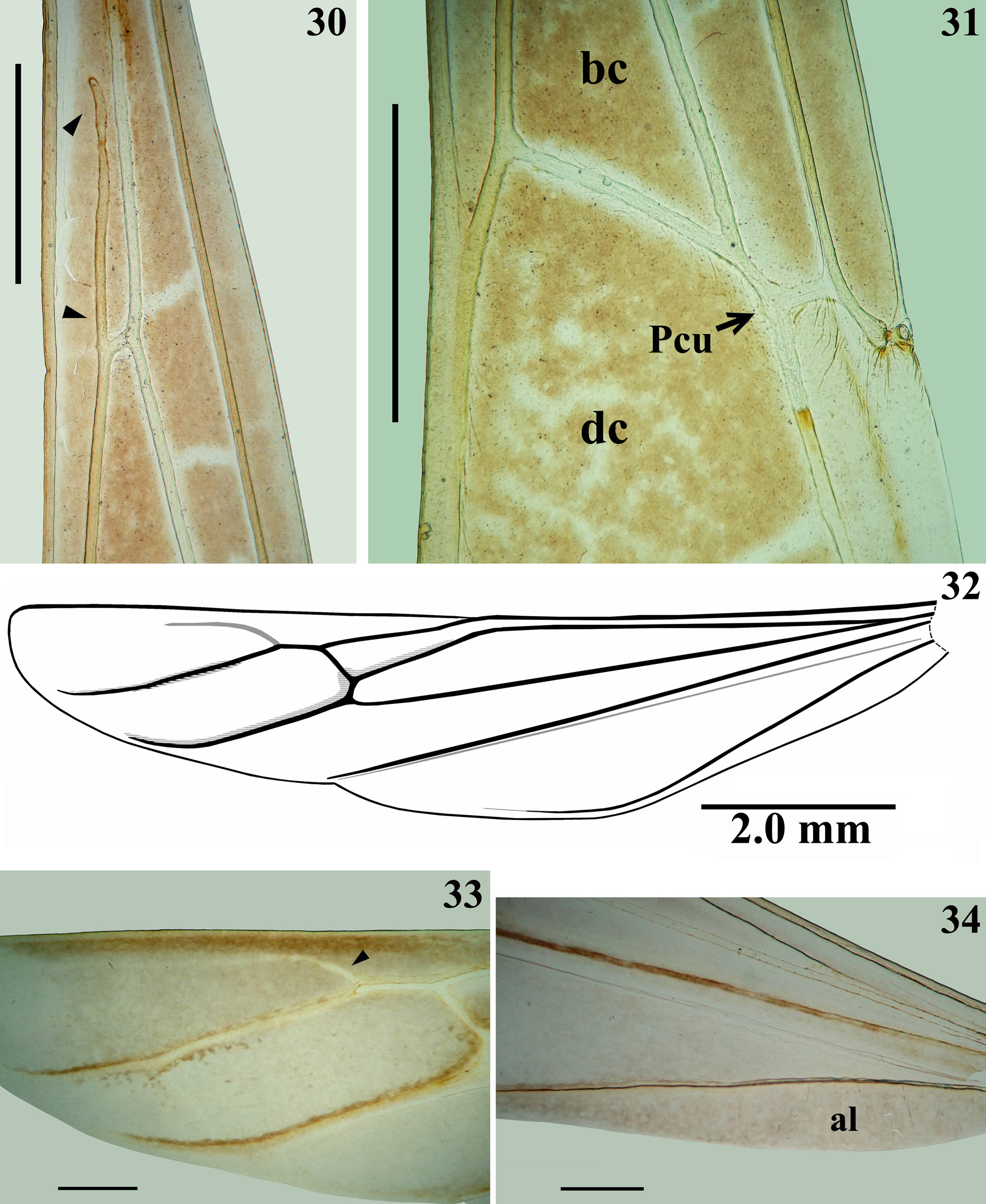
Structure. Integument mostly dull, moderately shiny, especially on legs. Head elongated; anteocular portion as long as postocular or approximately so. Transversal (interocular) sulcus deep, situated somewhat posteriorly to middle of eyes; just anterior to it, on midline, a small oval fossa and also on midline, posteriorly to this sulcus, an oval small shallow tumescence ( Figs. 2–3 View FIGURES 1 – 3 ). Eyes prominent, subcircular in dorsal view, reaching dorsal outline of head at interocular sulcus; not reaching ventral margin of head, which is near from inferior margin of the eye ( Figs. 2–3 View FIGURES 1 – 3 , 5 View FIGURES 4 – 8 ). Area just behind interocular sulcus somewhat elevated laterally ( Figs. 2–3 View FIGURES 1 – 3 , 5 View FIGURES 4 – 8 ). Dorsal interocular distance about twice or somewhat less than twice the width of each eye. Antenna inserted closer to apex of head than to the eyes ( Figs. 2–3 View FIGURES 1 – 3 , 5 View FIGURES 4 – 8 ); antennal segments progressively thinner, very slender; segments I and III straight; segment II somewhat curved at distal half; segment IV somewhat curved, with apex triangular ( Figs. 6–8 View FIGURES 4 – 8 , 9–18 View FIGURES 9 – 18 ). First two visible labial segments thicker than the distal segment; apex of segment III approximately at level of midportion of eye; segment IV ending at level of anterior extremity of stridulitrum ( Fig. 5 View FIGURES 4 – 8 ). Thorax: pronotum pedunculate; petiole approximately 1.4 to 1.6 times as long as fore lobe, the latter moderately widened; anterior projections of collar rounded; hind lobe bell-shaped in dorsal view, hind border emarginated at mid portion; humeral angle with an elevated rounded tubercle ( Figs. 1–2 View FIGURES 1 – 3 , 4–5 View FIGURES 4 – 8 ). Lateral margins of stridulitrum conspicuous. Spine of scutellum somewhat elongated, directed straight upwards; spine of metanotum smaller than the spine of scutellum and directed obliquely backwards ( Figs. 19–20 View FIGURES 19 – 23 ). Fore legs slender ( Figs. 4 View FIGURES 4 – 8 , 21 View FIGURES 19 – 23 ); fore coxae elongated, approximately two times as long as fore lobe of pronotum; fore tibiae shorter than fore femora, somewhat curved ( Figs. 4 View FIGURES 4 – 8 , 21 View FIGURES 19 – 23 ); somewhat depressed in dorsal portion subapically; thickened at apex, where the inner surface is somewhat flattened and has a very small subdistal pecten ( Fig. 23 View FIGURES 19 – 23 ). Mid and hind legs very long and slender, with tibiae thinner and longer than femora and slightly curved ( Figs. 24–26 View FIGURES 24 – 26 ). Tarsi short, three-segmented, slender ( Fig. 23 View FIGURES 19 – 23 ). Forewings ( Figs. 27–29 View FIGURES 27 – 29 ) slender, long, surpassing apex of abdomen by approximately 0.3 to 0.5 mm, venation as in Fig. 29 View FIGURES 27 – 29 ; basal cell subtriangular, with its base truncated and with two basally veins emitted from its base; the outer vein shorter ( Figs. 27–30 View FIGURES 27 – 29 View FIGURES 30 – 34 ); Pcu meeting discal cell somewhat beyond apex of basal cell ( Figs. 27–29 View FIGURES 27 – 29 , 31 View FIGURES 30 – 34 ); discal cell subpentagonal; pterostigma ending far from apex of the wing ( Figs. 27–29 View FIGURES 27 – 29 ). Hind wings reaching basal third of tergite VII; venation as in Fig. 32 View FIGURES 30 – 34 . Although there is a pale linear marking above R+M, which could suggest a branch from this vein, a microscopic examination of the region made it clear that it is not a true vein, and therefore R+M is not bifurcate ( Fig. 33 View FIGURES 30 – 34 ). Anal lobe with region between 2A and hind border not sclerotized, just somewhat pigmented ( Fig. 34 View FIGURES 30 – 34 ). Abdomen: segments II–III narrow; segments III–V progressively slightly widened towards apex; segment VI slightly narrowed towards apex; segment VII strongly narrowed towards apex, dorsally with a short tongue-shaped prolongation posteriorly, with a rounded posterior margin, slightly approaching tip of pygophore (35–36). Eight sternite covering approximately half of the pygophore, ventrally ( Figs. 37–38 View FIGURES 35 – 40 ). Connexival segments simple, straight ( Fig. 35 View FIGURES 35 – 40 ).
Male genitalia: pygophore subrectangular in dorsal and ventral views ( Figs. 38, 40–41 View FIGURES 35 – 40 View FIGURES 41 – 48 ); anterior dorsal sclerotization narrow; basal margin curved backwards on midportion ventrally ( Fig. 40–41 View FIGURES 35 – 40 View FIGURES 41 – 48 ); posterior process somewhat large, apex triangular, acute ( Fig. 39 View FIGURES 35 – 40 , 41–42 View FIGURES 41 – 48 ). Parameres symmetrical, curved and with moderately long, thin and non-numerous setae in distal half; apex rounded ( Figs. 38–40 View FIGURES 35 – 40 , 43 View FIGURES 41 – 48 ). Phallus symmetrical ( Figs. 44–45 View FIGURES 41 – 48 ). Articulatory apparatus ( art app) with basal plate extension elongated ( Figs. 44, 46 View FIGURES 41 – 48 ), basal plates fused at basal half and at apical portion ( Fig. 46 View FIGURES 41 – 48 ); basal arms ( ba) very short ( Figs. 44–46 View FIGURES 41 – 48 ). A pair of elongated ventral sclerotizations on phallosoma ( vsc) (= ventral sclerotization of phallobase sensu Wygodzinsky 1966), in shape of slender rods ( Figs. 44–45, 47 View FIGURES 41 – 48 ). Struts ( str) largely fused, widened toward apex, largely divided at basal extremity, and very narrowly at apical third (“incised” sensu Wygodzinsky 1966), between the oval-shaped lobes present at apical half ( Fig. 48 View FIGURES 41 – 48 ). Laterally to these lobes, and fused to them medially, weak somewhat large sclerotizations are present ( Fig. 48 View FIGURES 41 – 48 ). Between the ventral sclerotizations on the phallosoma ( vsc) and the struts ( str), three small and illdefined sclerotized, irregular thickenings, without forming clear processes, in approximately distal half of phallosoma ( Fig. 44 View FIGURES 41 – 48 ). Endosoma wall smooth, simple, without noticeable expansions.
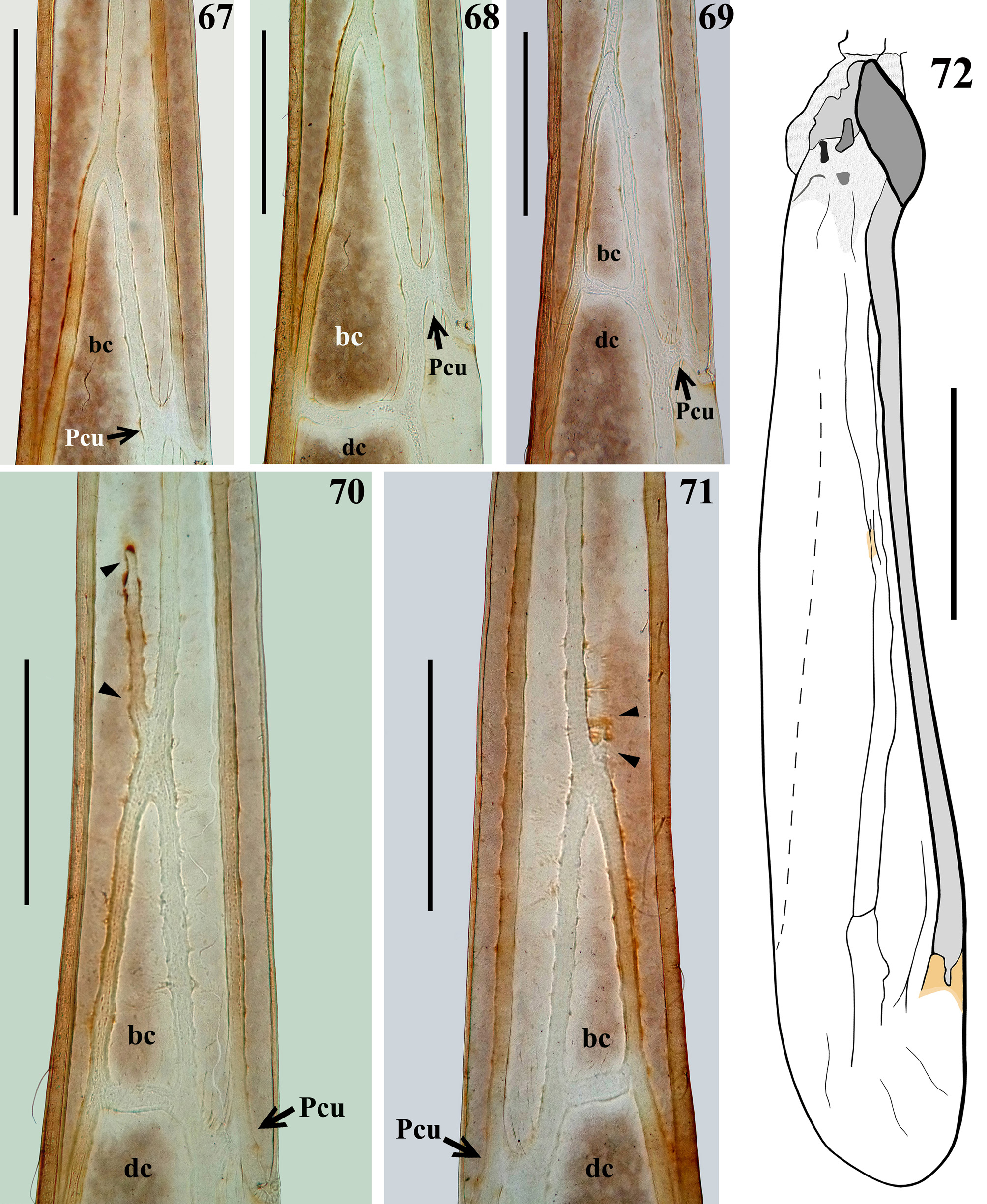
Female. Figures 49–79 View FIGURES 49 – 51 View FIGURES 52 – 54 View FIGURES 55 – 59 View FIGURES 60 – 63 View FIGURES 67 – 72 View FIGURES 73 – 79 . Measurements: Table 2. The following differences in relation to males were recorded. Coloration: Antenna: segment I: the first (subbasal) annulus approximately four times longer than the width of the segment, the next three annuli two to three times as long as the width of the segment and the fifth (subapical) annulus generally shorter than the width of segment ( Fig. 56 View FIGURES 55 – 59 ); segment II: three narrow pale annuli approximately twice to five times as long as the width of the segment ( Fig. 57 View FIGURES 55 – 59 ). Thorax: pale coloration on margins of meso- and hind metathoracic lobes somewhat broader. Whitish spots on posterior portion of mesopleura and metapleura somewhat larger. Legs: at apical portion of fore coxa, a moderately irregular large distal pale spot, interrupted on dorsal and ventral surfaces, and therefore not forming a ring ( Fig. 60 View FIGURES 60 – 63 ). Paler coloration on dorsal surface of fore femora restricted to approximately the proximal three-fourths; there are four pale spots along the segment, which are somewhat larger, reaching a greater extent on dorsal surface, and are also paler than those in males, the two additional spots lie between the midpoint and the whitish large apical spot, and sometimes they are united on inner surface, forming a longer spot; most or all of the large and medium-sized processes of both series with their basal portions pale to whitish, and the area surrounding their insertion generally pale too ( Fig. 60 View FIGURES 60 – 63 ). The markings, somewhat distal to midpoint on fore tibiae, are paler and sometimes form a complete smaller annulus ( Fig. 60 View FIGURES 60 – 63 ). Forewing mostly brownish; a transverse to oblique small, sometimes incomplete, pale stripe at middle portion of discal cell and a few small pale spots inside this cell and distally to posterior vein of the same; distal fourth with a moderately large oblique whitish stripe followed by a blackish apical portion (Figs. 64, 66). Hind wings hyaline with a small subapical reddish spot on upper margin and an even smaller, almost imperceptible reddish spot near middle portion of upper half of the wing ( Fig. 72 View FIGURES 67 – 72 ). Abdomen ( Figs. 73–77 View FIGURES 73 – 79 ) somewhat paler on tergite III and basal half of tergite IV, extensively marked with pale and whitish markings on segments IV–VII. Tergites IV–VII with a conspicuous scattered irregular pattern of pale and grayish markings. Connexivum with conspicuous whitish markings, on dorsal connexival segments: basal and median spots on segment III, large markings on basal and distal portions of segments IV–VI and subbasally and distally on segment VII; on ventral connexival segments: similar markings to the dorsal segments, but their extent is somewhat different: only a basal spot on segment III, a larger basal spot on segment IV; shorter on distal portions of segments IV–V, larger on basal portions of segments V–VI; on segment VII, the proximal spot is basally located. On the sternites, the whitish markings are generally more conspicuous than in male; sternite VII with an additional pair of submedian spots on basal half and another pair of sublateral spots on distal half. Genital segments darkened with pale markings on midportion of tergites VIII and IX; on the latter, more extensively and including the posterior margin too; on basomedian portion of gonocoxites. Vestiture: Antenna: segment I with a few longer thin scattered somewhat curved setae on inner surface at apical extremity too; about twelve isolated elements interpreted as trichobothria were observed on segment II. Fore femur: posteroventral series composed of about eleven to sixteen large and medium-sized spiniferous processes, among these, approximately seventy-one to ninety-one small spiniferous processes. Lengths of larger processes combined with apical spines about four-fifths of diameter of segment ( Fig. 61 View FIGURES 60 – 63 ). Anteroventral series composed of nineteen to twenty spiniferous tubercles, among which there are about one hundred and six to one hundred and thirtyfive sharp small spines. The setae accompanying the anteroventral series are somewhat larger. Ventral surface of fore tibiae with about 26 to 29 nine larger spiniferous processes. Structure. Head ( Figs. 52, 54–55 View FIGURES 52 – 54 View FIGURES 55 – 59 ): eyes smaller, inferior margin of the eye somewhat far from the ventral margin of head; area just behind interocular sulcus somewhat less elevated laterally; dorsal interocular distance about thrice the width of each eye. The small oval fossa anterior to midpoint of transverse sulcus somewhat more evident; oval shallow tumescence posterior to transverse sulcus smaller. Thorax: petiole somewhat more than twice to approximately 2.5 times as long as fore lobe; hind lobe proportionally smaller with hind margin somewhat narrower ( Figs. 52–53 View FIGURES 52 – 54 ). Spines of scutellum and metanotum obliquely directed upwards ( Fig. 59 View FIGURES 55 – 59 ). Fore coxae approximately 2.5 times as long as fore lobe of pronotum. Forewing narrow, short, reaching only the level of midportion to the distal third of tergite IV ( Figs. 49–51 View FIGURES 49 – 51 , 64–66, 73–74). Basal cell triangular, varying in size, approximately one-third in length, with a single directed vein emitted from its base (Figs. 64–69); in one paratype, however, in the left forewing, there is a short free vein emitted from the base of the basal cell, in addition to elongated vein leading to axillary region ( Fig. 70 View FIGURES 67 – 72 ), while in its right forewing there is a rudimentary basal portion of a similar short free vein ( Fig. 71 View FIGURES 67 – 72 ). Pcu variably positioned: ranging from meeting basal cell far above its apex to meeting discal cell near or somewhat farther beyond apex of basal cell (Figs. 64–71). Discal cell subrectangular (Figs. 64–66). Pterostigma ending relatively farther from apex of the wing (Fig. 65). Hind wing very short, reaching only the anterior border of tergite I (Fig. 66); its upper margin enlarged and somewhat sclerotized; at the end of this enlarged margin, a small area with a reddish tinge; membrane very thin with a few incomplete delicate apparent veins ( Fig. 72 View FIGURES 67 – 72 ). Abdomen: segments II–III narrow; segment IV widened towards apex, segments V– VI greatly widened; segment VII smaller and narrowed towards apex, dorsally with apical margin rounded and not covering genital segments ( Figs. 73–77 View FIGURES 73 – 79 ). The connexival segments are straight or slightly curved; the lateral margins of the dorsal plates are entire, while the postero-lateral angles of the ventral plates of segments IV–VI are very slightly expanded outwards ( Figs. 73–76 View FIGURES 73 – 79 ). The more enlarged portion of abdomen, from apical half of segment IV to apex of segment VI, on dorsal and ventral views is subrectangular in shape ( Figs. 73–76 View FIGURES 73 – 79 ); this portion, together with segment VII, forms a rounded bulge on lateral view while the pleural membrane is much more developed, folded and longitudinally pleated, in which folds have sclerotized strips ( Fig. 77 View FIGURES 73 – 79 ). External female genitalia ( Figs. 78–79 View FIGURES 73 – 79 ): simple; tergites VIII and IX forming a subhorizontal surface; tergite VIII moderately short, transversely subrectangular; tergite IX large, slightly narrowing towards apex, gonocoxites (of segment VIII) large, apex rounded.
Distribution. Brazil, state of Minas Gerais.
Etymology. The specific epithet is the combination of two indigenous words (Tupi-Guarani): kuara, meaning “hole”, “nest”, “hiding place” and jucassaba which means “killer”, “killing” referring to the condition of a cave hunter. To be treated as a noun in apposition, ending not to be changed.
Specimens examined. BRAZIL, Minas Gerais, Pains, MALE HOLOTYPE: Gruta da Torre [“tower cave”] II, ( 20º25'21.04"S 45º36'03.58"W), 03.ii.2017, R.L. Ferreira leg. ( MNRJ) GoogleMaps ; PARATYPES: Gruta da Torre [“tower cave”] II, ( 20º25'15.92"S 45º36'08.08"W), R.L. Ferreira leg., 03.ii.2017, 1 male, 2 females ( ISLA 21277) GoogleMaps ; 01.x.2015, 1 female ( MNRJ), 2 females ( ISLA 12095, 12081); Gruta do Ronco [“snoring cave”], ( 20º25'50"S 45º36'41"W), 28.xi.1999, R.L. Ferreira & M.S. Silva leg., 1 male ( ISLA 12086). GoogleMaps
Habitat and natural history. Specimens of D. kuarajucassaba sp. nov. were found in three caves in the karst region of Arcos-Pains-Doresópolis (state of Minas Gerais, Brazil) ( Figs. 80–81 View FIGURES 80 – 83 ). Specifically, the caves where the specimens were found are located in the municipality of Pains. This region presents the largest concentration of caves in South America (around 2,500 known caves). More than 300 caves were sampled in the area, and specimens were only found in three caves. However, it is interesting to note that these caves present similar conditions, including small dimensions (less than 100 meters) ( Fig. 81 View FIGURES 80 – 83 ), presence of dry substrates and entrances with many collapsed blocks ( Fig. 80 View FIGURES 80 – 83 ). Furthermore, the caves are located in a specific place in the southern region of the karst area, near the urban area of the municipality of Córrego Fundo.
The specimens were always found on the cave walls, usually near the ceiling ( Figs. 82–83 View FIGURES 80 – 83 ). Most of them were found near the cave entrances, but always in disphotic areas. These regions usually present many accidental or transient invertebrate species; thus, it is plausible to assume that specimens of D. kuarajucassaba sp. nov. prefer these areas because of the regular supply of prey. Nymphs were also observed in the caves, indicating that the species might be reproducing in such habitats, what would indicate a troglophilic condition. However, since we did not perform any collections in the epigean surroundings, it is impossible to determine the importance of the caves as a habitat for the species. Furthermore, the area has undergone intense impact from mining activities (since the 1960’s), which remove the limestone to produce cement. In addition, the natural vegetation of the area has also suffered intense changes over recent decades, especially due to expansion of agricultural activities. This has resulted in a rather fragmented landscape, in which the remaining vegetation is mainly associated with the tops and immediate surroundings of the limestone outcrops (which are unsuitable for human use). Accordingly, it is also possible that the species has been sheltering in caves due to the severe alterations to which the external habitats have been subjected over recent years. Thus, further studies focusing on this species are essential to determine what their current distribution is (in epigean habitats), as well as whether they are threatened.
Comments. Almost all the features of the male of D. kuarajucassaba sp. nov. are in accordance with the definition of Dohrnemesa ( Wygodzinsky 1945, 1966, Forero 2004), therefore the placement of D. kuarajucassaba sp. nov. in this genus is justified.
The only discordances relate to its total length (18.5–18.8 mm), which is somewhat longer than the length of the species previously attributed to Dohrnemesa ( 10–16 mm); and to the region between 2A and the hind border of the anal lobe in the hind wing ( Fig. 34 View FIGURES 30 – 34 ), which is not sclerotized in D. kuarajucassaba sp. nov., although it has been recorded as distinctly sclerotized in Dohrnemesa ( Wygodzinsky 1966).
All the features of the male genitalia of D. kuarajucassaba sp. nov. ( Figs. 37–48 View FIGURES 35 – 40 View FIGURES 41 – 48 ) are very similar to those of the other species of Dohrnemesa too, as summarized by Wygodzinsky (1966).
The male of D. kuarajucassaba sp. nov. most closely resembles the male of D. lanei Wygodzinsky, 1945 . This species was described based on a single male from the state of São Paulo, Brazil ( Wygodzinsky 1945), with no further records of it.
The males of these species share similarities, including the total length (18.5–18.8 mm for D. kuarajucassaba sp. nov. and 16.0 mm for D. lanei ), general dark coloration with extensive pale markings distributed in a similar pattern on thorax and legs, and many structural features. The few features of the male genitalia in D. lanei that have been described ( Wygodzinsky 1945), such as shape and general appearance of the posterior process of pygophore and parameres, seem very similar in D. kuarajucassaba sp. nov. too. The two species can be separated by the following set of characteristics: 1—antennal segments I and II with five and three pale annuli respectively in D. kuarajucassaba sp. nov. ( Figs. 6–7 View FIGURES 4 – 8 ) and four pale annuli in both segments in D. lanei ; 2—fore femora with a pair of pale spots on basal half, and an apical larger whitish spot in D. kuarajucassaba sp. nov. ( Fig. 21 View FIGURES 19 – 23 ) and 4–5 whitish annuli in D. lanei ; 3—sternites with several whitish markings in D. kuarajucassaba sp. nov. ( Fig. 36 View FIGURES 35 – 40 ) and uniformly blackish in D. lanei ; 4—postocular portion of head with a median distinct tubercle ( D. lanei ) or an oval small shallow tumescence ( D. kuarajucassaba sp. nov.); 5—lengths of larger processes of fore femora combined with apical spines about three-fourths of diameter of segment ( D. kuarajucassaba sp. nov.) or about as long as its diameter ( D. lanei ); 6—spine of scutellum straight ( D. kuarajucassaba sp. nov., Figs. 4 View FIGURES 4 – 8 , 19–20 View FIGURES 19 – 23 ) or obliquely directed upwards ( D. lanei ); 7—Pcu meeting discal cell somewhat distad ( D. kuarajucassaba sp. nov., Figs. 27–29 View FIGURES 27 – 29 , 31 View FIGURES 30 – 34 ) or proximad ( D. lanei ) of the apex of basal cell on forewing; 8—connexival segments simple, straight, in D. kuarajucassaba sp. nov. ( Fig. 35 View FIGURES 35 – 40 ), and somewhat enlarged at posterolateral angles in D. lanei .
An attempt was made to locate the holotype of D. lanei . Wygodzinsky (1945) mentioned that it had been deposited in the collection of the Zoology Sector of the Department of Agriculture of the State of São Paulo. This institution and its collection were incorporated by the University of São Paulo, into its Zoology Museum (“Museu de Zoologia da Universidade de São Paulo ”, MZSP)( Marinoni & Marinoni 2012). However, after a careful search in the collection of this institution, Dr Carlos Campaner, of the curator team of MZSP, kindly informed the first author that the holotype of D. lanei was not found there. Therefore, it can be considered to have been lost.
Dohrnemesa lanei View in CoL was considered to be close to D. santosi Wygodzinsky, 1945 View in CoL , which was also described based on a single male from the state of São Paulo ( Wygodzinsky 1945, 1966). The holotype of D. santosi View in CoL remains deposited in the National Museum of Rio de Janeiro (MNRJ) (Figs. 84–87), with no other known specimens of this species. It is noteworthy that some of the features pointed out by Wygodzinsky (1945, 1966) for separating D. lanei View in CoL from D. santosi View in CoL are shared by the latter and D. kuarajucassaba View in CoL sp. nov. such as: connexival segments simple, straight and with pale markings; Pcu meeting discal cell somewhat distad of the apex of discal cell in forewing; larger processes of fore femora combined with apical spines shorter than diameter of segment.
However, the general coloration pattern of D. santosi View in CoL (Fig. 84) is quite diverse from D. lanei View in CoL and D. kuarajucassaba View in CoL sp. nov. ( Fig. 1 View FIGURES 1 – 3 ). Additionally, D. kuarajucassaba View in CoL sp. nov. and D. lanei View in CoL have a petiole that is distinctly longer than the fore lobe of the pronotum and the process of the pygophore is acute and triangular, while in D. santosi View in CoL the petiole is not longer than the fore lobe (Fig. 84) and the process of pygophore is rounded. There is no tubercle or elevation at the median portion of the postocular region in the latter species, either ( Wygodzinsky 1945, 1966).
Surprisingly, some features of the female of D. kuarajucassaba sp. nov., particularly its brachypterous condition, are in accordance with the definition of Polauchenia , such as: forewings greatly reduced, reaching about middle of abdomen ( Figs. 49–51 View FIGURES 49 – 51 , 64, 74) and with a single directed vein emitted from the base of their basal cell (Figs. 64–66); extremely reduced hind wings (Fig. 66); abdomen wider ( Wygodzinsky 1966).
Nevertheless, two differences were promptly recorded. The total length (21.1–23.0 mm) is slightly longer than previously recognized for Polauchenia ( 11–20 mm in length) ( Wygodzinsky 1966) and the abdomen is not lobate.
Considering that there was only a small difference in length, this can possibly be accepted as a small extension of the range of total length in Polauchenia , thus increasing it from 11–20 mm to 11–23 mm.
Concerning the shape of the [enlarged portion of the] abdomen, although Wygodzinsky (1966) considered it always to be lobate in brachypterous females of Polauchenia , his assumption was based only on the female of P. marcapata Wygodzinsky, 1966 , the sole species in which a brachypterous female had been known until now. Therefore, for the taxonomic value of the lobate abdomen to be ascertained, the presence or absence of this feature would need to be determined among the females of more species.
On the other hand, the recorded variation of the venation of the forewings in females of D. kuarajucassaba sp. nov. deserves some comments. The Pcu was found to be variably positioned, ranging from meeting the basal cell far proximad of its apex to meeting the discal cell somewhat farther distad of the apex of the basal cell (Figs. 64–71).
Although in most of the females the basal cell had a single directed vein emitted from its base, in one female paratype, the left forewing clearly showed a short free vein emitted from the base of the basal cell in addition to an elongated vein leading to the axillary region ( Fig. 70 View FIGURES 67 – 72 ), while its right forewing showed a rudimentary similar short free vein ( Fig. 71 View FIGURES 67 – 72 ). At first sight, it would be plausible to consider these variations to be simple isolated anomalies, especially regarding the wing reduction in the present case. However, it is noteworthy that the presence of this short vein emitted from the base of the basal cell is one of the main differences among the few that exist between Polauchenia and Dohrnemesa, genera that are considered close to each other ( Wygodzinsky 1945, 1966), while the position at which the Pcu met the basal cell was considered to have taxonomic significance in these genera, for separating the species ( Wygodzinsky 1966). The venation of the wings is regarded as furnishing excellent taxonomic characteristics at the generic and tribal levels in Emesinae ( Wygodzinsky 1966) and has been used extensively to diagnose and/or separate supra-specific taxa, without questioning their possible intra-specific variation ( Gil-Santana & Marques 2005). Gil- Santana et al. (1999) recorded intra-specific variation in the number of cells in the forewing of Mayemesa lapinhaensis ( Wygodzinsky, 1950) . Because the number of cells in the forewing was the main characteristic separating Mayemesa from another genus of Emesini , the monotypic genus Amilcaria, Gil-Santana et al. (1999) proposed that Amilcaria should be regarded as a junior synonymy of Mayemesa . Taking into account that most species of Dohrnemesa and Polauchenia have been described and/or recorded based on a single or only a few specimens ( McAtee & Malloch 1925, Wygodzinsky 1945, 1947, 1950, 1958, 1966, Maldonado 1968, Forero 2006, Gil-Santana & Ferreira 2016), the possible intra-specific variation of some characteristics may remain unknown. Therefore, the significance of the abovementioned variations in the forewings of females of D. kuarajucassaba sp. nov., i.e., whether they are merely isolated anomalies associated with a brachypterous condition, or whether they are truly intra-specific variations with possible taxonomical inferences, will be ascertained if or when more specimens of both Dohrnemesa spp. and Polauchenia spp. are examined in the future.
Additionally, the female of D. kuarajucassaba sp. nov. shows a general resemblance to the female of Polauchenia marcapata . This species was described based on two females from Peru ( Wygodzinsky 1966). On the basis of Wygodzinsky (1966), Forero (2006) referred that P. marcapata would also have been known from Bolivia. However, Wygodzinsky (1966) listed his type specimens as being from Marcapata River ( holotype) and Valle Chanchamayo ( paratype), both located in Peru. Maldonado (1990) also cited the species as being limited to Peru. Forero (2004) recorded this species in Colombia, but subsequently ( Forero 2006) alleged that the identification of the specimen from Colombia, which he had considered to be very close to P. marcapata , required confirmation because the humeral angles of the pronotum were pointed rather than blunt, and the color pattern of the hemelytra was less marked than in P. marcapata . Hence, additional specimens would be needed to reaffirm the identification. Therefore, so far, only Peru and possibly Colombia should be included in the geographic distribution of P. marcapata .
The following structural differences can promptly separate the females of D. kuarajucassaba sp. nov. and P. marcapata , respectively: 1—length: 21.1–23.0 mm versus 17.5 mm; 2—postocular region of the head: without pointed elevations versus with distinctive pointed elevations; 3—shape of posterior widening of abdomen: subrectangular versus subspherical; and 4—posterior margin of connexival segments: not lobate versus lobate.
Nonetheless, the decision to make the current placement of D. kuarajucassaba sp. nov. in Dohrnemesa was taken primarily because of the features of its male, which were shown to be very compatible with those attributed to Dohrnemesa ( Wygodzinsky 1945, 1966, Forero 2004).
Although the females of D. kuarajucassaba sp. nov. present similarities to P. marcapata rather than to known females of species of Dohrnemesa, this does not compromise the current placement of the new species in the latter genus.
This is firstly because in half of the species of Dohrnemesa, only males are known ( Wygodzinsky 1945, 1958, 1966, Gil-Santana & Ferreira 2016), including in D. lanei , which was considered to be close to D. kuarajucassaba sp. nov. It is possible that some (unknown) females of other species of Dohrnemesa have features similar to those of D. kuarajucassaba sp. nov., but have remained unrecorded so far. Additionally, with the exception of the apterous species D. feminata , for which placement in Dohrnemesa is uncertain, the known females of Dohrnemesa spp. ( D. buyassuana Wygodzinsky, 1958 , D. carvalhoi Wygodzinsky, 1966 and D. reimoseri Wygodzinsky, 1950 ) are fully winged. Therefore, because D. kuarajucassaba sp. nov. is the first species of Dohrnemesa with a brachypterous female that has been discovered, features associated with reduced wings have only been recorded from one female of this genus so far. Hence, some apparent similarities between D. kuarajucassaba sp. nov. and P. marcapata may be more related to their brachypterous condition than to systematic proximity between them.
Secondly, it is possible that the venation in reduced forewings of females is prone to anomalies, which would diminish or impair their use as reliable taxonomic characteristics. In this case, absence of a short free vein emitted from the base of the basal cell should be disregarded as a reliable generic characteristic for differentiating between the two genera under discussion in relation to brachypterous females. In this regard, the assumption of Wygodzinsky (1966) that the venation in the reduced forewing of brachypterous females of Polauchenia would be complete can be shown not to be true, taking into account what was recorded here in relation to females of D. kuarajucassaba sp. nov.
Thirdly, considering that no males of P. marcapata have yet been described, it is possible that the taxonomic position of this species might need to be reviewed if or when males are reported.
Fourthly, given that Polauchenia and Dohrnemesa seem to be closely related genera ( Wygodzinsky 1945, 1966), occurrence of variable degrees of resemblance between the females of species of these genera could be quite possible.
Regarding sexual dimorphism in Dohrnemesa spp., a universal sexual difference among Emesinae is the width of abdomen, which is generally slight in fully winged forms, but sometimes very conspicuous if the female has reduced or absent wings ( Wygodzinsky 1966). In fact, while the abdomen was recorded as somewhat less wide at the middle portion in the male of Dohrnemesa carvalhoi , compared with its female ( Gil-Santana & Ferreira 2016), the females of D. kuarajucassaba sp. nov. present abdominal segments IV–VI that are much wider ( Figs. 49–51 View FIGURES 49 – 51 , 73– 77 View FIGURES 73 – 79 ) than those of the males (4, 35–36). In the latter species, the eyes of the males were shown to be evidently larger ( Figs. 2–3 View FIGURES 1 – 3 , 5 View FIGURES 4 – 8 , 52, 54–55 View FIGURES 52 – 54 View FIGURES 55 – 59 ), while those of the males of D. carvalhoi were only somewhat larger ( Gil-Santana & Ferreira 2016), like in many other species of Emesinae , in which the eyes of the females are smaller than those of the males ( Wygodzinsky 1966). Females of D. kuarajucassaba sp. nov. were also recorded as being larger and more conspicuously marked, mainly on enlarged abdominal segments. The latter difference was particularly more pronounced on the exposed tergites (i.e. not covered by the shortened forewing) ( Figs. 35 View FIGURES 35 – 40 , 73–75 View FIGURES 73 – 79 ).
On the other hand, the remarkable development of the pleural membrane in the enlarged portion of the abdomen (distal half of segment IV and subsequent segments) in the females of D. kuarajucassaba sp. nov., which was folded with longitudinal sclerotized strips ( Fig. 77 View FIGURES 73 – 79 ), is striking. While this resembles the abdomen of females of the triatomine Dipetalogaster maximus (Uhler, 1894) , in which an analogous folded and longitudinally pleated abdominal pleura allows unusual expansion of the abdomen when the specimen feeds ( Lent & Wygodzinsky 1979), no similar description could be found among species of Emesinae .
The discovery of D. kuarajucassaba sp. nov. makes it necessary to make slight modifications of the detailed generic description of Dohrnemesa provided by Wygodzinsky (1966): total body length is 10–16 mm for other species, but reaches 18.8 (males)–23 (females) mm in D. kuarajucassaba sp. nov.; region between 2A and hind border of the anal lobe in the hind wing sclerotized or not; females fully winged or brachypterous; brachypterous females show: forewings with variable venation, reaching at most distal third of tergite IV; hind wing very short, reaching anterior border of tergite I; abdominal segments IV–VI enlarged.











| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Emesinae |
|
Tribe |
Emesini |
|
Genus |
Dohrnemesa kuarajucassaba
| Gil-Santana, Hélcio R. & Ferreira, Rodrigo Lopes 2017 |
D. kuarajucassaba
| Gil-Santana & Ferreira 2017 |
D. kuarajucassaba
| Gil-Santana & Ferreira 2017 |
D. kuarajucassaba
| Gil-Santana & Ferreira 2017 |
Dohrnemesa lanei
| Wygodzinsky 1945 |
D. santosi
| Wygodzinsky 1945 |
D. santosi
| Wygodzinsky 1945 |
D. lanei
| Wygodzinsky 1945 |
D. santosi
| Wygodzinsky 1945 |
D. santosi
| Wygodzinsky 1945 |
D. lanei
| Wygodzinsky 1945 |
D. lanei
| Wygodzinsky 1945 |
D. santosi
| Wygodzinsky 1945 |