
Conochironomus tobaterdecimus (Kikuchi & Sasa)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4109.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:CFD3FDAC-B477-4607-9FB7-251DCDB479E9 |
|
DOI |
https://doi.org/10.5281/zenodo.6068469 |
|
persistent identifier |
https://treatment.plazi.org/id/37607C47-FF8F-5F45-A290-CCBFFB42F91E |
|
treatment provided by |
Plazi |
|
scientific name |
Conochironomus tobaterdecimus (Kikuchi & Sasa) |
| status |
|
Conochironomus tobaterdecimus (Kikuchi & Sasa) View in CoL
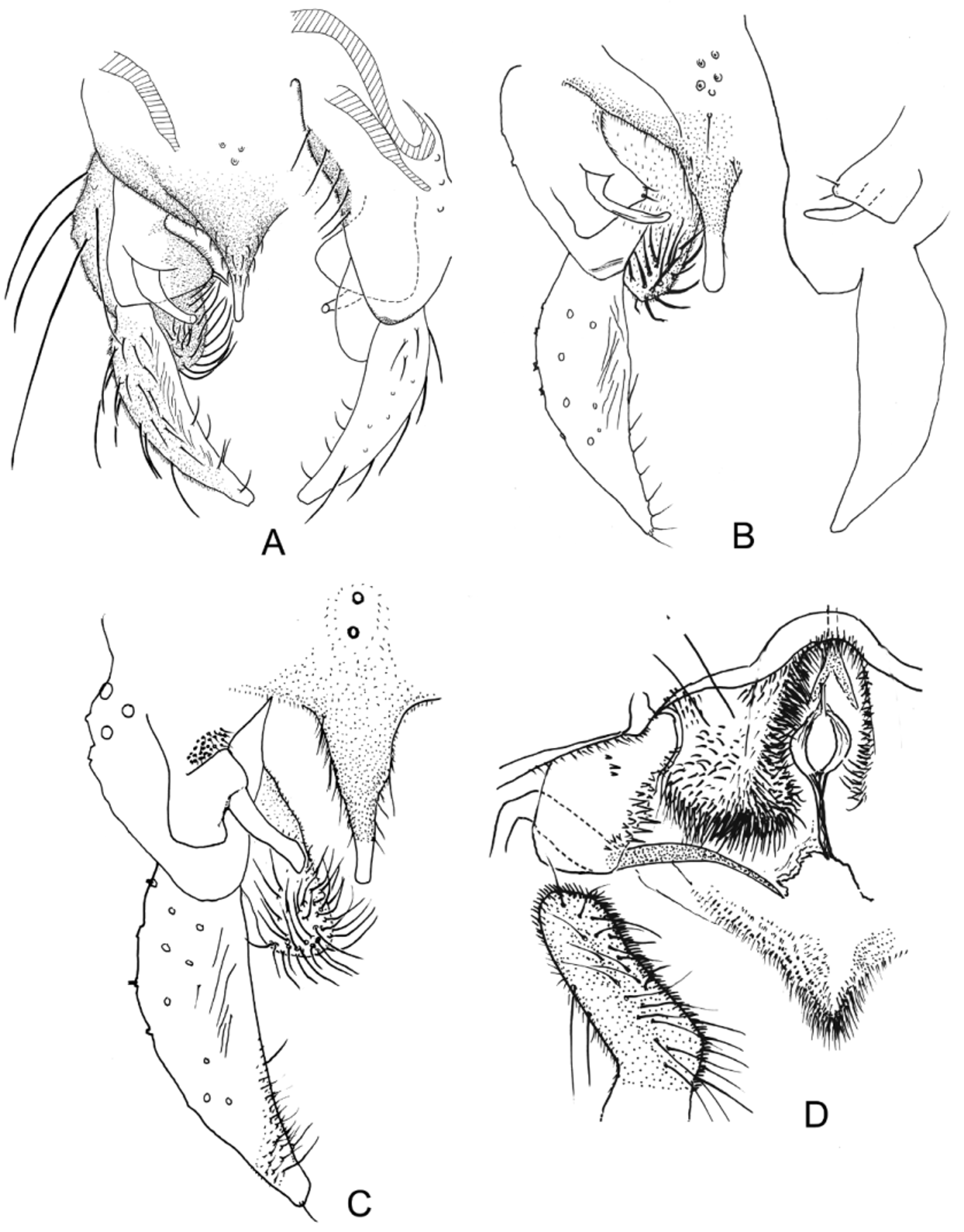
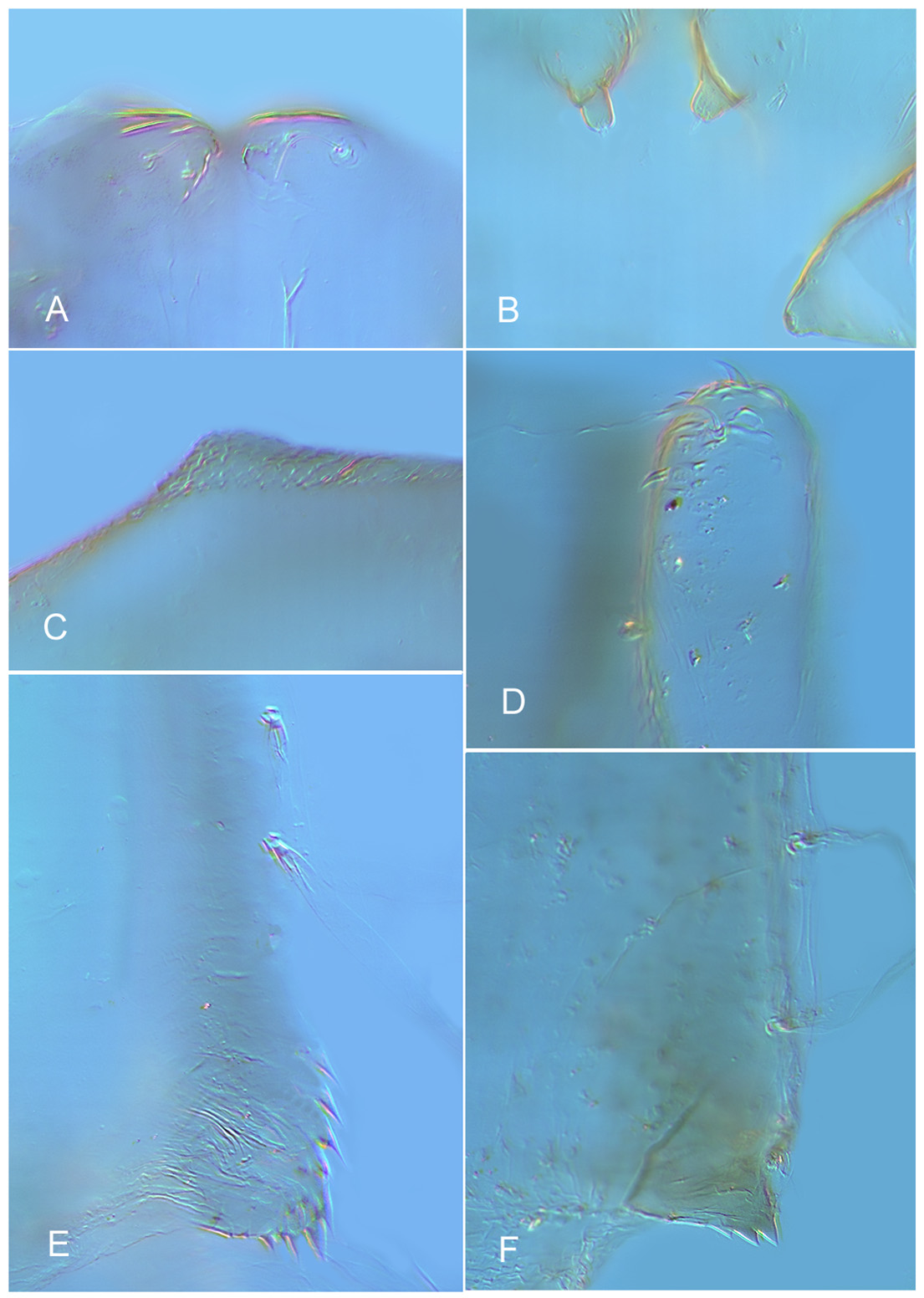
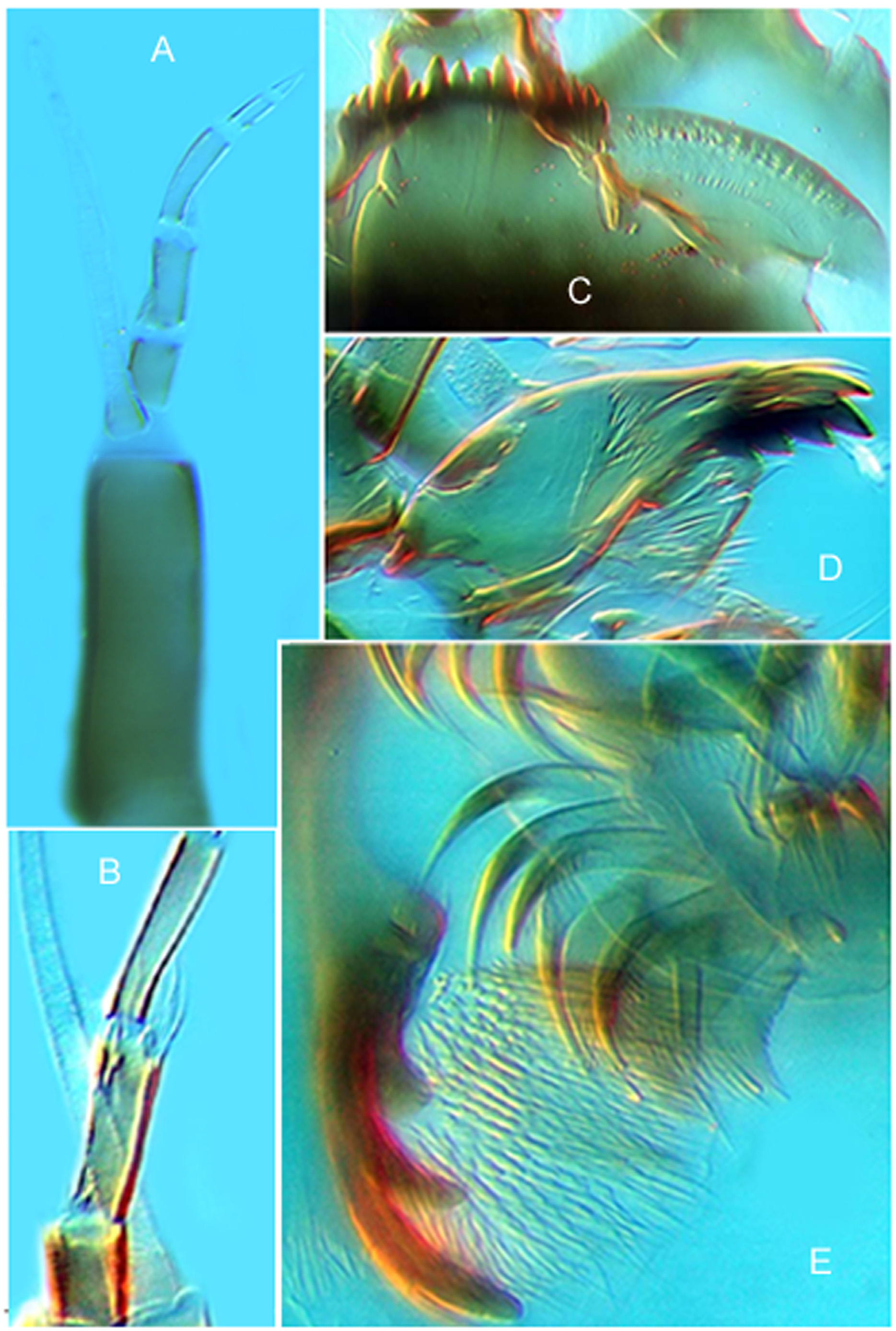
( Figs. 1 View FIGURE 1 A–E; 2C,D; 3E,F; 4A,C; 5A–C, D, F)
Sumatendipes tobaterdecimus Kikuchi & Sasa, 1990 ( Kikuchi & Sasa 1990: 313) View in CoL .
Chironomus (Conochironomus) View in CoL " tokunagai " of Karunakaran (1969), unavailable name ( ICZN 1999: Article 8a). Conochironomus tobaterdecimus ( Kikuchi & Sasa, 1990) View in CoL . Comb. n. (merely inferred in Cranston 2004: 715).
Material examined. All slide mounted in Euparal; SINGAPORE: ♂, Bukit Timah N.P., Jungle Falls, 1°21’21”N 103°48’26”E, 12.iii.2009 (Cranston); ♂, Bedok Reservoir, Floating deck A, 1°20’N 103°55’E 13.iv.2013, emergence trap, CP379 ( TMSI team) (GenBank KU507300 View Materials ); ♀, Upper Seletar Reservoir, forest area, 1°24’10”N 103°48’27”E, emergence trap, 16.vii.2013, CP459 ( TMSI team) (association by barcode, GenBank 507304); 4L, THAILAND: Roi Et Prov., Chaturaphak Phiman District, Nong Lad, 15°53'36"N 103°32'54"E, 1.iii.2012 (Simwisat) (association by barcode).
Putative immature material. Pupae. Pe, SINGAPORE: Bedok Reservoir, NE shore, 1°20’47”N 103°55’31”E, 23.ii.2012 (Ang); Larvae. L, SINGAPORE: Central Catchment, Nee Soon Swamp, 1°23'N 103°48’E, 13.iii.2009 (Cranston); L, same as preceding except 27.ii.2012 (MV NS 27-2-12 #1).
Redescription (partial; additional material substantially conforms to previous descriptions of all stages in Karunakaran 1969, sub Chironomus (Chironomus) tokunagai ; of male in Kikuchi & Sasa 1990, sub Sumatendipes tobaterdecimus ).
MALE ( Figs. 1 View FIGURE 1 A–D; 2C; 3E, F) (n = 1–2). Body dark brown with slightly darker delimited thoracic vittae, and paler pronotum, trochanters, femoral apices and distal sections of tibiae ( Fig.1 View FIGURE 1 A, B). Mensural features as in Table 1 View TABLE 1 . Genitalia ( Figs. 1 View FIGURE 1 D; 2C; 3E, F) with few anal tergite setae located in mid-tergite anterior to base of elongate anal point flanked with lateral setae; anal point tapering to narrower parallel-sided medial section, narrowly rounded at apex. Superior volsella structurally complex ( Figs. 2 View FIGURE 2 C, 3E), basal lobe bearing strong microtrichia on median (inner) contour, otherwise smooth; sinuous digitiform projection arises from broad base dorsally on basal lobe, initially dorsally directed, then narrowed and curved medio-posteriorly, terminating in up-turned, rounded tip; Basal lobe without strong setae, digitiform projection bare. Median volsella absent. Inferior volsella basally fused to medial margin of gonocoxite, with swollen free apex densely covered with thick, recurved, simple setae, none directed posteriorly. Gonostylus tapering, with weak inner creases ( Fig. 3 View FIGURE 3 F), terminating in rounded apex.
FEMALE ( Figs. 1 View FIGURE 1 E; 2D) (n = 1). Body length c 5.4 mm. Antenna: flagellomeres 1–4 500 µm, terminal (5th) 300 µm; AR 0.6. Thoracic setation: ac absent, dc 7, pa 3, sct 6. Wing length 3.0 mm, numbers of setae on wing veins R 34, R1 34, R4+5 40, on sq 14. Genitalia ( Figs. 1 View FIGURE 1 E; 2D). Notum thin, long, extending full length of segment, flared posteriorly prior to short rami. Seminal capsules oval, abruptly darkened in distal 1/3 to base of very short neck; spermathecal ducts straight, broad, ending separately. Gonapophysis VIII in ventral view ( Figs. 1 View FIGURE 1 E, 2D) clearly divided into large, quadrate, densely chaetose dorsomesal lobe and slightly smaller, rectangular ventrolateral lobe bearing spine-like chaetae on its median submargin, otherwise essentially bare. Cerci elongate rectangular in dorsal view.
PUPA ( Figs. 4 View FIGURE 4 A,C; 5A–C), based on tentatively associated exuviae (n = 1). Length c. 8 mm, pale, with brownish margins to thoracic appendages, abdominal segment apophyses indistinct.
Cephalothorax. Cephalic tubercle ( Fig. 4 View FIGURE 4 A) squat, 12 µm high, with hyaline but strong frontal seta, 50µm. Pedicel sheath with one inner tubercle. Antepronotum dorsally tuberculose, with 1 hyaline dorsal seta; l.apn not visible. Dorsal region of scutum weakly creased, non-rugose; scutal tubercle with tuberculose surface ( Fig. 4 View FIGURE 4 C). Thoracic horn hyaline; number of branches not detectable in slide preparation; tracheal bundle simple, ovate. No prealar tubercle.
Abdomen. Tergal armament as in Fig. 5 View FIGURE 5 A. Segments I and II without spinules. Hook row comprising 55 hooks in uniserial row, extending c. 60% of width of tergite II. Tergite III with wide and deep area of spinules, this area smaller and ending more anteriorly on T IV and V, T VI with anterior transverse band only, T VII without spinules; T VIII with antero-medial transverse patch, T IX with wide patch of spinules. Conjunctives bare. Most sternites bare, S VII and VIII with large quadrate area of spinules ( Fig. 5 View FIGURE 5 B). Caudolateral corner of segment VIII ventrally with 4–5 transversely aligned, basally fused, straight spines ( Fig. 5 View FIGURE 5 C). Pedes spurii B strong on segment II, absent on III. Pedes spurii A (vortices) absent. Segments V–VIII with 0, 0, 2, 5 taeniate lateral setae. Anal lobe dorsally with broad spinulose area ( Fig. 5 View FIGURE 5 A, B) with multiserial fringe of c. 100 taeniae (not shown), without dorsal seta.
LARVA ( Fig. 5 View FIGURE 5 , D, F; 6A–E) (n = 4). Conforms to generic diagnosis ( Cranston & Hare 1995) and closely resembles Australian C. australiensis . Body length c. 7–9 mm.
Head capsule with dark postoccipital margin; most of postmentum and posterior 1/3 of head darkened, anteriorly paler yellow-brown. Eye double, with larger spot exactly dorsal to smaller ventral spot. Body red, claws golden to golden brown (posterior). Head capsule length c. 490–530, postmentum length 164–180. Dorsal head and frontoclypeal apotome as in Cranston & Hare (1995: fig. 7h); Antenna ( Fig. 5 View FIGURE 5 D; Cranston & Hare 1995: fig 7e) with segment lengths (base to apex): 68–75; 13–15; 18–22; 8–12; 5–7. AR 0.9–1.2; Lauterborn organs large, alternate, 8–9 long; style slender, 10 long; blade 78–82 long, extending to apical segment or slightly beyond. Mandible (identical to Fig. 6 View FIGURE 6 D; Cranston & Hare 1995: fig. 7f) 155–165, with somewhat darkened outer tooth as long as dark apical tooth; two dark inner teeth; mola includes small darkened distal area close to base of long, simple seta subdentalis. Labrum (as in Cranston & Hare 1995: fig. 7g) with SI setae arising from common, fused bases (illustrated but not stated in Cranston & Hare 1995); SI and SII finely plumose; pecten epipharyngis of 3 separated scales, each with 2–3 blunt teeth; premandible 87–92, with 3 well-developed teeth and small basal 4th tooth. Mentum ( Fig. 5 View FIGURE 5 F; Cranston & Hare 1995: fig. 7a–d) total width 130–152, with 4 median (ventromental) teeth, varying in relative height of median pair of teeth (1 specimen has only 3 teeth; Pramual et al. 2016: fig. 4), and varying in pigment intensity from yellow-brown to as dark as lateral (dorsomental) teeth. Ventromental plate ( Fig. 5 View FIGURE 5 F) 62–65 apart medially, single plate 162–178 long, with characteristic ultrastructure ( Cranston & Hare 1995: fig. 7b) and variably wavy anterior margin.
Abdomen. Anterior parapod claws simple, forming dense cluster. Procercus and supraanal setae pale-mid brown.
Remarks. Type material of Sumatendipes tobaterdecimus was not examined. Recognition, including membership of Conochironomus , is based on the description and drawings of Kikuchi & Sasa (1990), plus images of the holotype male genitalia available at http://www. type.kahaku.go.jp/TypeDB (species name misspelled 'tobaterdecumus'). The anal point of the type appears to differ in shape from Singaporean males, but this may arise from the poor preparation and distortion of the specimen (as too often in material from Sasa’s studies).
Identical DNA barcoding COI sequences allow association of an adult male and female from Singapore (above). Larvae of Conochironomus collected from Nee Soon ( Singapore) provided no DNA capable of allowing further association; thus, larvae (and pupae) are associated only putatively with C. tobaterdecimus . Later five larval specimens (CP461 from Bedok reservoir; CP1136, 1137, 1716, 1245 from Upper Seletar Reservoir) yielded barcodes identical to C. tobaterdecimus (GenBank accessions KU507299 View Materials , KU507301 View Materials -3, KU507305 View Materials , respectively) but have not been examined by the author. Two larvae sequenced for barcode COI by Pramual et al. (2016, GenBank codes KT213039 View Materials and KT213040 View Materials ) are less than 1.5% different from adult C. tobaterdecimus from Singapore. This value lies well within the values of 4–5% taken to reflect species differences in Tanytarsus ( Lin et al. 2015) and found appropriate in Cricotopus ( Krosch et al. 2015) ; this boundary may have applicability across all Chironomidae . Unfortunately vouchers were not retained (P. Pramual, pers. comm. 2015) and morphology had to be derived from photographs of menta. However, further specimens from the same locality have been provided for morphometrics and barcoding, confirming association with C. tobaterdecimus from Singapore.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Conochironomus tobaterdecimus (Kikuchi & Sasa)
| Cranston, Peter S. 2016 |
Chironomus (Conochironomus)
| Cranston 2004: 715 |
Sumatendipes tobaterdecimus Kikuchi & Sasa, 1990 ( Kikuchi & Sasa 1990: 313 )
| Kikuchi 1990: 313 |