
Paradoneis carmelitensis, Arriaga-Hernández, Stefan, Hernández-Alcántara, Pablo & Solís-Weiss, Vivianne, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3686.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:DBCCE386-C7F3-4AA6-B233-AC3E44D53A19 |
|
DOI |
https://doi.org/10.5281/zenodo.5672327 |
|
persistent identifier |
https://treatment.plazi.org/id/3750D77E-DF0F-087A-FF4F-8A71FB81EC5C |
|
treatment provided by |
Plazi |
|
scientific name |
Paradoneis carmelitensis |
| status |
sp. nov. |
Paradoneis carmelitensis View in CoL n. sp.
Figures 4 View FIGURE 4 A–C, 5A–I
Material examined. Type material. Holotype: National Polychaete Collection, ICML, UNAM (CNP– ICML POH– 02–002): Station 24, 18° 34' 30”N, 91° 29' 54”W, 3.9 m. Paratypes: National Polychaete Collection, ICML, UNAM (CNP– ICML POP–02–002): 9 paratypes, one of them coated with gold for SEM studies: Station 24, 18° 34' 30”N, 91° 29' 54”W, 3.9 m. Museo Nacional de Ciencias Naturales de Madrid ( MNCN 16.01/14720): 6 paratypes, Station 24, 18° 34' 30”N, 91° 29' 54”W, 3.9 m. Stations located in Términos Lagoon, Campeche, Southern Gulf of Mexico; collected October 20, 2009, by P. Hernández-Alcántara.
Additional material. National Polychaete Collection, ICML, UNAM (CNP– ICML PO–02–041), 42 specimens: Station 16, 3 specimens, March 13, 2009, 18° 29’ 23”N, 91° 33’ 30”W, 3.5 m. Station 18, 2 specimens, March 13, 2009, 18° 33’ 40”N, 91° 36’ 10”W, 4 m. Station 18, 1 specimen, October 20, 2009, 18° 33' 40”N, 91° 36' 10”W, 4 m. Station 22, 8 specimens, October 20, 2009, 18° 39.40.40’ 30”N, 91° 32’ 44”W, 2.6 m. Station 23, 3 specimens, October 20, 2009, 18° 37’ 00”N, 91° 31’ 10”W, 3.9 m. Station 24, 25 specimens, October 20, 2009, 18° 34’ 30”N, 91° 29’ 54”W, 3.9 m. All the stations located in Términos Lagoon, Campeche, Southern Gulf of Mexico; collected by P. Hernández-Alcántara.
Description. Holotype complete with 86 chaetigers, body length 10.51 mm, width 0.23 mm. Paratypes with 42–88 chaetigers, 5.54–13.02 mm long and 0.13–0.26 mm wide. Body long, nearly cylindrical, slightly flattened dorsally in branchial region ( Figs 4 View FIGURE 4 A, 5A). Prostomium conical, longer than wide, with an apical lobe retracted at the tip ( Fig. 5 View FIGURE 5 D). Eyespots absent. Two ciliary bands limited to dorsal surface on anteriormost region of the prostomium, posterior band emerging laterally from nuchal slits and circumscribing ventral surface ( Figs 4 View FIGURE 4 A, 5D). Ciliary bands also present on dorsum of prebranchial and branchial segments ( Fig. 5 View FIGURE 5 D). Peristomium well developed, fused ventrally to chaetiger 1, forming lower margin of mouth. A pair of nearly longitudinal nuchal organs as deep nuchal slits, in posterior half of prostomium. First three notopodial postchaetal lobes (prebranchial chaetigers) short, conical-triangular shaped ( Figs 4 View FIGURE 4 A, 5C); from chaetiger 4 (first branchial) they become longer, digitiform ( Figs 4 View FIGURE 4 A, 5E), and then slightly shorter, triangular in postbranchial region; they become longer again at 9 to 11 chaetigers before the pygidium ( Figs 4 View FIGURE 4 C, 5H). Without postchaetal lobes. Branchiae from chaetiger 4 ( Figs 4 View FIGURE 4 A, 5A); 12 pairs, cylindrical, basally wider, distally rounded, longer than body width. Postbranchial segments bito triannulated. Notopodial modified lyriform chaetae from first branchial segment (chaetiger 4) down to 17 (in some paratypes to chaetigers 12–16), 1–2 chaetae per parapodium ( Fig. 5 View FIGURE 5 F, G). Both tines of lyriform chaetae similarly thick, but one filament is clearly longer with thin tip ( Fig. 4 View FIGURE 4 B); both filaments with spinulate inner margins ( Fig. 5 View FIGURE 5 G). All other chaetae are long capillaries, hirsute along one margin ( Fig. 5 View FIGURE 5 B), with a long tip. In the last 3–4 chaetigers, the number of chaetae decreases to 1–3 per ramus ( Figs 4 View FIGURE 4 C, 5H), with one simple capillary chaeta stouter than the other accompanying simple chaetae. Pygidium rounded with two long mid-lateral cirri and one short mid-ventral cirrus ( Figs 4 View FIGURE 4 C, 5H, I).
Remarks. The Genus Paradoneis was erected by Hartman (1965) to include paraonids with notopodial modified chaetae (lyriform chaetae or simple spines) and no antenna. Nowadays, this genus is represented by 19 species, including the new species described in this study, and two subspecies ( Aguirrezabalaga & Gil 2009; De León-González & Díaz-Castañeda 2011). Paradoneis lyra (Southern) had been previously recorded in the lagoonestuarine systems from the southern Gulf of Mexico ( Hernández-Alcántara & Solís-Weiss 1991); however, its record is questionable, since the species was first described from shallow waters of Ireland (Ballynaki Harbour, Galway) ( Southern 1914). Later, this species was widely reported around the world and erroneously considered cosmopolitan, since many of the more detailed descriptions of P. l y r a were recognized as belonging to other species ( Mackie 1991).
On the other hand, Aguirrezabalaga and Gil (2009) analyzed the Paraonidae from the Bay of Biscay, northeastern Atlantic, describing eight new species (two of which belong to the genus Paradoneis ), and made an exhaustive compilation of the diagnostic features from each Paradoneis species that had been described until then. After that, only Paradoneis strelzovi from the eastern Pacific has been described ( De León-González & Díaz- Castañeda 2011).
According to the detailed information supplied by Aguirrezabalaga and Gil (2009), there are only five species of Paradoneis with three prebranchial chaetigers whose modified chaetae are exclusively lyriform on notopodia: P. brunnea (Hartmann-Schröder & Rosenfeldt) from the Antarctic Ocean, P. forticirrata (Strelzov) recorded from the Pacific Ocean, P. l y r a (Southern) from Ireland, P. nipponica Imajima from Japan and P. perdidoensis (McLelland & Gaston) recorded in Florida. Although the segment where the lyriform chaetae starts is variable, P. carmelitensis n. sp. from Términos Lagoon ( Table 2 View TABLE 2 ) can be separated from P. perdidoensis , P. nipponica and P. brunnea , because in all these species the modified chaetae (lyriform) always appear initially on prebranchial chaetigers (chaetigers 2 or 3), but especially because P. perdidoensis bears only 3-4 pairs of branchiae, and P. nipponica has the highest number of branchiae among the Paradoneis species (23-26 pairs); P. brunnea bears a similar number of branchial pairs (12), but unlike P. carmelitensis n. sp., their branchiae are distinctly shorter, less than the segment’s width. Finally, P. forticirrata can be differentiated from P. carmelitensis n. sp., because of a greater number of branchial pairs are present (15–17), but mainly because its notopodial lobes in prebranchial segments are massive ( Strelzov 1973).
In this sense, P. carmelitensis n. sp., seems only close to P. l y r a. However, diagnostic characters such as distribution of the lyriform notochaetae, number of branchiae and prebranchial chaetigers, shape and size of the postchaetal lobes and the presence of neuropodial hooks in posterior chaetigers, described from specimens collected far away from its type locality ( Ireland), do not necessarily correspond with the original description of P. lyra ( Mackie 1991) . Thus, when we analyzed in detail the morphology of the specimens collected in Términos Lagoon, the differences observed stand out, such as the typical subdermal brown eyes (sometimes absent) of P. l y r a which are always lacking in P. carmelitensis n. sp.; in addition, the postchaetal lobes of the prebranchial region are digitiform in P. l y r a, while in P. carmelitensis n. sp., they are triangular (wide basally). Besides, in the specimens from Términos Lagoon the branchiae are longer than the distance between the branchial bases.
Branchial length Longer than body width
Notopodial lobes of prebranchial, branchial and Short, conical-triangular in prebranchial chaetigers; longer, postbranchial segments digitiform from chaetiger 4, becoming slightly shorter, triangular in postbranchial region, but lengthening again in chaetigers near the pygidium
Shape of notochaetae Forked (lyrate)
Modified notochaetae (chaetiger) 4 to 17 (12–16)
Acicular neurochaetae (chaetiger) No
Anal cirri 3: two longest mid-lateral; one short mid-ventral Number of chaetigers>82
Etymology. The name carmelitensis is derived from the gentilitious "Carmelita", name given to the people living in the area of Términos Lagoon.
Habitat. In mud and fine sands, depth 2.6 to 4 m, temperature 25.7°C to 28.09°C, salinity 31.74 to 35.85 psu, pH 7.8 to 9.1.
Distribution. Center and southern regions of Términos Lagoon, southern Gulf of Mexico.
Spatial and seasonal distribution of Paraonidae . The new species of paraonids, Aricidea (Acmira) hirsuta and Paradoneis carmelitensis , were collected only in eight stations (40%) from the 20 stations studied in Términos Lagoon. The paraonids were distributed mainly in the central and southern regions of the lagoon ( Fig. 1 View FIGURE 1 ), representing only 0.9% (5 specimens) of the 527 polychaetes collected during the dry season but 43.5% (1178 specimens) of the 2708 polychaetes collected during the rainy season.
The spatial and seasonal distribution at the species level is also noteworthy since, during the dry season, only specimens from P. carmelitensis n. sp., were collected and only in two stations located at the southwestern end of the lagoon (Sta. 16 and 18), while during the rainy season both species were recorded, mainly in seven stations from its central and southern regions ( Figs 1 View FIGURE 1 , 6 View FIGURE 6 ). During the rainy season, the increased number of the paraonids was due almost exclusively to Aricidea (A.) hirsuta n. sp., which, with 1125 specimens, constituted 97% of the paraonids.
It is also interesting to note that station 18 (central region of the lagoon) was the only one where paraonids were found in both seasons, even though their densities were low (<20.7 ind./ 0.1 m ²): during the dry season only P. carmelitensis n. sp., was collected but during the rainy season both species were present ( Fig. 6 View FIGURE 6 ).
During the rainy season, each species displayed a different distribution pattern: Aricidea (A.) hirsuta n. sp., was found abundantly in the central region of the lagoon, whereas P. carmelitensis n. sp., was restricted to its central-oriental region. Only in three stations from the central region both species were present simultaneously: 22 (204.7 ind./ 0.1 m ²), 23 (586.8 ind./ 0.1 m ²) and 24 (232.1 ind./ 0.1 m ²) ( Fig. 6 View FIGURE 6 ); in these stations the highest densities of Paraonidae were also found. On the contrary, in the southern and western areas, the occurrence of paraonids decreased drastically and only Aricidea (A.) hirsuta n. sp., was collected with very low densities (1.7 to 2.5 ind./ 0.1 m ²).
Although P. carmelitensis n. sp., was the only species collected in both seasons, it was also the less abundant paraonid (mean = 6.01 ind./ 0.1 m ²) and its distribution was not linked significantly to any of the environmental parameters measured. On the contrary, Aricidea (A.) hirsuta n. sp., was only found during the rainy season, but its abundance variations were directly correlated to changes in salinity (r² =0.74, p <0.05) and temperature (r² =0.71, p <0.05). That is, in this study, the greater abundance of Aricidea (A.) hirsuta n. sp., in the middle lagoon was associated with salinities of 35.38 ups and temperatures of 27.8°C, on average.
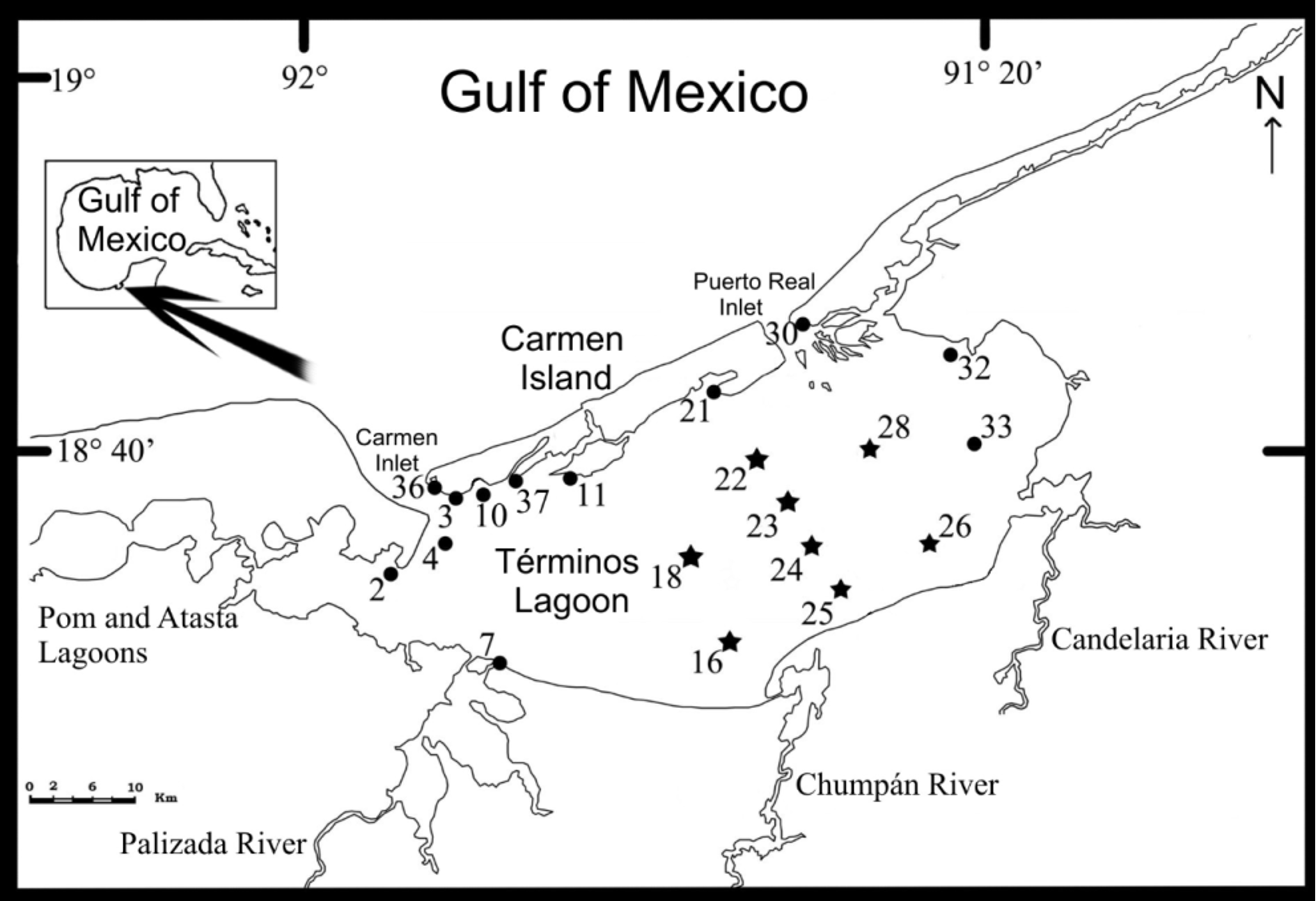
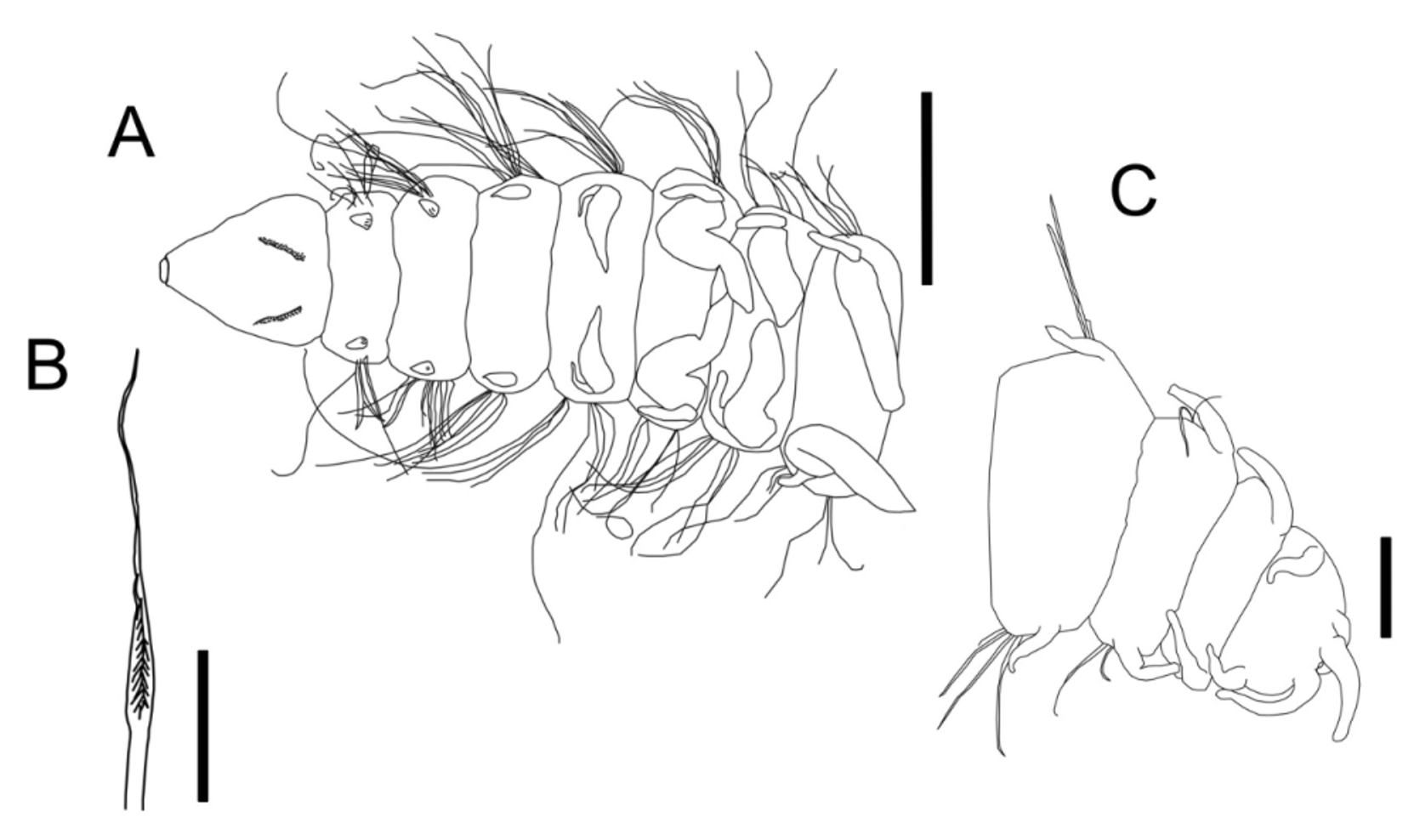

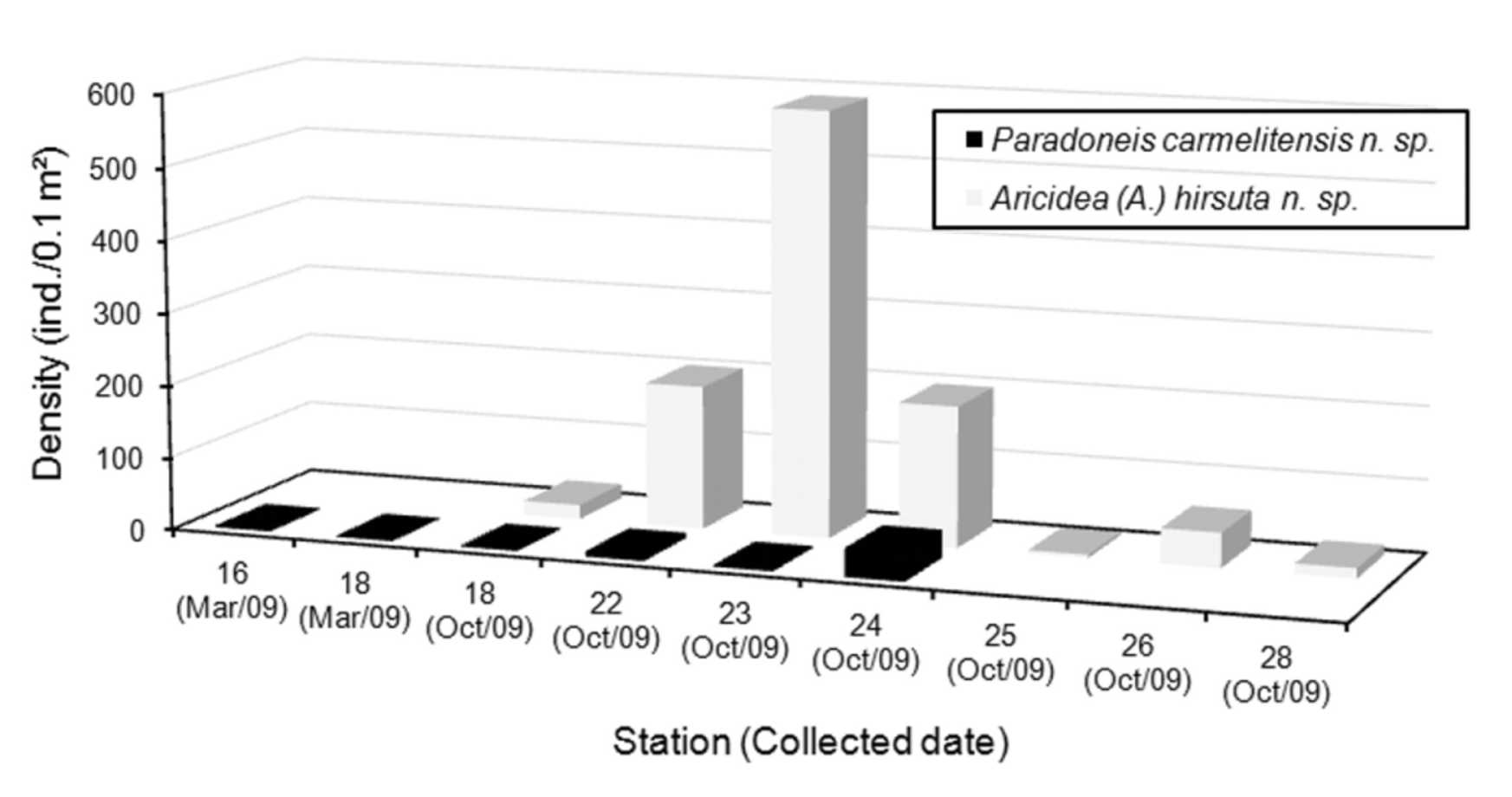
TABLE 2. Diagnostic characters of Paradoneis carmelitensis n. sp. based on Aguirrezabalaga and Gil (2009).
| Diagnostic characters | Paradoneis | carmelitensis n. sp. |
|---|---|---|
| Prebranchial chaetiger | 3 | |
| Branchial pairs | 10 to 13 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |