
Microtelphusa furcifer, Pedraza, Manuel & Tavares, Marcos, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3847.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:665B2C62-758D-489B-8A20-228D48EDCEC3 |
|
DOI |
https://doi.org/10.5281/zenodo.5672242 |
|
persistent identifier |
https://treatment.plazi.org/id/361D878B-FFFD-FFA5-FF62-FE2A9431F93B |
|
treatment provided by |
Plazi |
|
scientific name |
Microtelphusa furcifer |
| status |
sp. nov. |
Microtelphusa furcifer n. sp.
( Figs. 1A–E View FIGURE 1. A – E ; 2A–C; 3A–D)
Type material. Holotype: male cl 16.01, cw 25.03 mm, Guyana, Potaro-Siparuni Region, Kuribrong River, 5º22'35''N, 59º33'09''W, approximately 440 meters altitude, M.C. Folane et al. coll., 23-24 March 2011 ( MZUSP 32309). Paratypes: 1 male cl 15,49 mm, cw 24.41 mm ( INPA 2051), 1 male cl 16.69 mm, cw 25.12 mm ( MZUSP 24494), same data as holotype.
Comparative material. Microthelphusa barinensis : male holotype, Venezuela, Barinas, 24 February 1968, G. Rodríguez coll. and leg. ( IVIC 643); 2 males, 3 females paratypes ( IVIC 643), same data. Microthelphusa bolivari : male holotype, Venezuela, Bolivar, El Dorado-Santa Helena road, altitude 1350 m, 24 October 1972, E. Sierra coll., G. Rodríguez leg. ( IVIC 644); 1 male, Bolívar, El Dorado-Santa Helena road, Km. 126, altitude 1500 m, 16 April 1973, L. Andreani coll., G. Rodríguez leg. ( MHNLS 865). Microthelphusa forcarti : 6 males, 1 female, Venezuela, Mérida, Tabay, Acequia Creek, altitude 1710m, 21 October 1970, G. Rodríguez coll., G. Rodríguez leg. ( IVIC not numbered). Microthelphusa lipkei : male holotype, Brazil, Amazonas state, Barcelos, Aracá mountain, Anta Creek, 00°54'40''N, 63°26'19'' W, altitude 1133 m, 03‒09 August 2007, M. Moreira & J.R. Andrade do Rosário coll., C. Magalhães leg. ( INPA 1748). Microthelphusa meansi : male holotype, Guyana, Potaro Siparuni, Wokomung Massif, middle camp, 5º06'36''N, 59º 49'14'' W, altitude 1220 m, 18 July 2003, D. Means coll., N. Cumberlidge leg. ( USNM 1098372); 1 male paratype ( USNM 1098373), same data. Microthelphusa racenisi : male holotype, Aragua, Rancho Grande, 5 August 1949, S. Racenis coll., G. Rodríguez leg. ( IVIC 647); 10 males, 1 female, Venezuela, Aragua, Rancho Grande, Cloroní road, 23 March 1973, Fihn coll., G. Rodríguez leg. ( IVIC 648). Microthelphusa sucreensis : male holotype, Venezuela, Sucre, Elvecia, 15 January 1930, M. Netting coll., G. Rodríguez leg. ( USNM 230091); 3 males, 1female, Monagas, Negro River, close to Serrania, altitude 1980 m., 18 November 1993, A. Viloria coll., G. Rodríguez leg. ( IVIC 1087). Microthelphusa turumikiri : 5 males paratypes, Venezuela, Sucre, Turumiquiri hill, altitude 1500 m, 23 June 1968, A. Rivero coll., G. Rodríguez leg. ( IVIC 649). Microthelphusa viloriai : male holoype, Venezuela, Trujillo, Santa Ana Creek, altitude 1500m, 30 November 1989, J. Moscó et al. coll., H. Suárez leg. ( IVIC 1129); 5 males, 6 juveniles, Trujillo, El Rincón Creek, altitude 1550 m, 11 November 1991, A. Viloria et al. coll., H. Suárez leg. ( IVIC 1130). Microthelphusa wymanni : 1 male, Suriname, Brownsberg Nature Preserve Park, Witti Creek, 0 5 September 1977, R. Sawyer coll., M. Campos det. ( USNM 1155132); 1 male syntype, Suriname, M. J. Rathbun det. ( USNM 29975); 10 males, 7 females, 5 juveniles, Suriname, Brownsberg Nature Preserve Park, Mazoroni Creek, 100 Km from Paramaribo, 10 May 1977, G. Rodríguez leg. ( IVIC 1079).
Diagnosis. Apical plate plate of first gonopod distally produced into well-developed, wide, rounded lobe tapering abruptly laterodistally, ending in well-defined triangular lobe; mesial process deeply furcate; caudal and cephalic lobes nearly equal in length; caudal surface of apical plate provided with spinules; proximal spine-like outgrowth on mesial margin of apical plate absent; apical plate and mesial process nearly equal in size.
Description of holotype. Carapace dorsal surface smooth, moderately convex, regions barely defined; gastric pits almost obsolete. Cervical groove nearly straight, faint proximally, deep terminally, ending just before anterolateral margin. Postfrontal lobules small, poorly defined; median groove indistinct between postfrontal lobes; fronto-orbital region slightly bent anteriorly. Branchial region distinctly convex, covered with small rounded granules. Anterolateral margin fringed with small, anteriorly-directed tubercles almost evenly spaced. Posterolateral margin poorly defined, with minute sparse granules; molting suture well distinct. Front upper border clearly distinct, uninterrupted, fringed with minute granules; frontal margin carinate, strongly sinuous in frontal view, lined with minute papillae. Supra-, infraorbital margins lined with small tubercles; infraorbital margin strongly marginated, sloping mesialy; exorbital angle prominent, obtuse. Inner orbital tooth well developed, broad triangular, lobe-like, partially occluding orbital cavity. Subhepatic region with well spaced, small, rounded granules almost reaching to anterolateral margin of carapace. Pterystogomial regions covered by short pubescence. Mxp3 dactylus rather flattened dorsoventrally, concave ventrally; propodus, carpus subcylindric; carpus-propodus mesial, lateral margins heavily setate.
Mxp3 merus rimmed with distinctly carina, markedly shorter than ischium, about 0.71 times its length; posteromesial border with subquadrate, dentate, setate lobe; lobe grooved to receive folded dactylus. Mxp3 ischium visibly wider than merus; mesial edge setate, dentate terminally, teeth subquadrate, corneous proximally; lateral margin convex. Mxp3 exopod distinctly short, reaching to about 0,34 times length of lateral margin of ischium. Opening of efferent branchial channel large, devoided of setae; upper margin subcircular, complete; lower margin interrupted; channel closed ventrally by large lamellar scaphognathite. Epistome distinctly short, wide, deeply grooved transversally, upper margin straight, strongly carinate; lower margin deeply sinuous to receive folded mxp3 palp, broad triangular epistomial tooth in center filling gap between joining palps. Mandibular palp three-segmented, terminal segment inserted near mid-length of mesial surface of segment 2.
Major cheliped missing. Chelipeds noticeably heterochelous, similarly armed, right cheliped largest (as revealed by paratypes). Merus of minor cheliped subtriangular in cross-section, upper margin heavily crenulated; upper lateral surface scarcely squamose, lower lateral surface with indistinct low squamiform tubercles; ventromesial, ventrolateral margins toothed, teeth larger on ventromesial margin. Upper surface of carpus with low squamiform tubercles arranged in subparallel transversal ridges; mesial margin toothed, teeth increasing in size anteriorly. Propodus upper surface with low squamiform tubercles extending onto proximal dorsal surface of dactylus; lateral surface smooth in appearance, minutely punctate; mesial, ventral surfaces with sparse low squamiform tubercles; lateral, mesial surfaces of fixed, movable fingers; ventral, dorsal surfaces of fixed, movable fingers, respectively, with setigerous punctations arranged in longitudinal subparallel lines. Cutting edges of fingers with small, massive teeth; fingers gaping medially, proximally, closing terminally. P2‒P5 slender, P3 longest, P5 shortest; meri, carpi, propodi massive, dactili noticeably slender, with 5 longitudinal rows of strong acute spines: 3 dorsal, 2 ventral; propodi with minute scattered spines dorsally, ventrally; laterally punctate, mesially smooth; carpi minutely spinous dorsally, punctate laterally, smooth mesially, ventrally; meri crenulate dorsally, otherwise smooth.
Thoracic sternum rather longer than broad; sternal sutures 4/5 to 6/7 well marked, interrupted medially, ending just before reaching median line of thoracic sternum; sternal suture 7/8 well marked, uninterrupted; median line prolonging from sternite 4 to 8, deeper at interception with sternal suture 7/8; episternites 4‒6 triangular posteriorly, episternite 7 posteriorly truncate. Sterno-abdominal cavity deeply excavated, scarcely pubescent, pubescence rather long; tubercle of the abdominal holding system minute, placed near sternal suture 5/6.
Male abdomen with 6 somites, telson, triangular in shape, tapering progressively from third somite onward, margins scarcely pubescent, sutures between somites well distinct, complete; third somite widest, sixth longest with ventrolongitudinal ridge anterolaterally, ridge heavily setate. Telson broadly triangular, lateral margins distinctly crenulated. Penis noticeably long, emerging from nearby coxo-sternal condyle articulation, located in shallow depression on sternite 8, proximally thick, abruptly tapering distally.
G1 straight, columnar, almost as wide proximally as distally; large proximal semicircular process expanding mesially in cephalic view. Marginal suture rather straight, subparallel to longitudinal axis, slightly displaced toward mesial side in caudal view; row of long setae about midlength of caudal surface longitudinal axis between marginal, lateral sutures in caudal view. Lateral suture deep, extending 2/3 of gonopod length from proximal to terminal portions. Marginal lobe short, broadly triangular, separated from lateral surface by deep notch, slightly extending beyond spermatic channel. Field of dense apical spines, stretching diagonally to longitudinal axis, delimited laterally by caudal, cephalic lobes. Spermatic channel opening proximally in the field of apical spines, near convergence of marginal, cephalic lobes. Apical plate distally produced into a well-developed, wide, rounded lobe. Caudal, cephalic lobes nearly equal in length, separated by deep notch. Apical plate with distinct process tapering abruptly laterodistally ending in well-defined triangular lobe, slightly twisted to cephalic side apically in lateral view. Proximal spine-like outgrowth on mesial margin of apical plate absent. Caudal surface of apical plate provided spinous. Mesial process deeply furcate. G2 straight, almost as long as G1, conspicuously slender, distal 3/ 4 tapered; tip flattened, with numerous short spinules on sternal surface.
Type locality. Guyana, Potaro-Siparuni region, Kuribrong River, 5º22'35''N, 59º33'09''W, 440 meters of altitude.
Distribution. Presently known only from the type locality.
Etymology. From the Latin furcifer for "fork-bearing", in allusion to the morphology of the G1, which is characterised by a mesial process having a deeply furcate distal part.
Remarks. Microthelphusa furcifer n. sp. is herein assigned to the genus Microthelphusa , whose diagnostic characters (cf. Rodríguez, 1982 and Suárez, 2006) are readily recognized in the new species, namely, a mxp3 exopod that is about 1/3 the length of the ischium and a straight and short G1 having a field of apical spines stretching diagonally to the longitudinal axis of the gonopod. The mesial process of G1 is also large, flattened, distally triangular and the inferior margin lobular. Its mesial process is distinctly separated from the apical plate and the marginal lobe joins with the distal lobe.
Microthelphusa furcifer View in CoL n. sp., M. meansi View in CoL and M. rodriguezi View in CoL are unique in the genus in having a distinct laterodistal process emerging from the G1 apical plate, which is absent in all other presently known congeners. The new species can be readily separated from all congeners by having a G1 mesial process that is deeply furcate ( Figs. 2A, C View FIGURE 2. A – C ; 3A, C–D), whereas in M. meansi View in CoL the mesial process is broadly furcate and entire in the remaining congeners. Microthelphusa furcifer View in CoL n. sp. also differs from particular congeners by having the: (1) the distal margin of the process of the apical plate tapering abruptly laterodistally to end in a well-defined triangular lobe ( Figs. 2A–C View FIGURE 2. A – C ; 3A–D) (tapering progressively laterodistally to end in a poorly-defined, broad triangle in M. rodriguezi View in CoL ); (2) G1 apical plate distally produced into a well-developed, wide, rounded lobe ( Figs. 2A–C View FIGURE 2. A – C ; 3A–D) (narrow lobe in M. bolivari View in CoL , M. odaelkae View in CoL , M. sommani , M. turumikiri View in CoL , and M. wymani View in CoL ); (3) G1 caudal and cephalic lobes nearly equal in length ( Figs. 2B View FIGURE 2. A – C ; 3B) (distinctly unequal in length in M. barinensis View in CoL , M. bolivari View in CoL , M. forcarti View in CoL , M. ginesi View in CoL , M. odaelkae View in CoL , M. racenisi View in CoL , M. sucreensis View in CoL , M. turumikiri View in CoL , M. viloriai View in CoL and M. wymani View in CoL ); (4) caudal surface of the G1 apical plate provided with spinules ( Figs. 2A View FIGURE 2. A – C ; 3A) (spinules absent in M. barinensis View in CoL , M. bolivari View in CoL , M. forcarti View in CoL , M. ginesi View in CoL , M. racenisi View in CoL , M. sucreensis View in CoL , M. sommani , M. turumikiri View in CoL , M. viloriai View in CoL , M. turumikiri View in CoL , and M. wymani View in CoL ); (5) absence of the proximal spine-like outgrowth on the mesial margin of the G1 apical plate ( Figs. 2A View FIGURE 2. A – C ; 3A) (spine-like outgrowth present in M. barinensis View in CoL , M. bolivari View in CoL , M. lipkei View in CoL , M. sommani , and M. wymani View in CoL ); (6) the G1 apical plate and mesial process nearly equal in size ( Figs. 2A, C View FIGURE 2. A – C ; 3A, D) (distinctly unequal in size in M. barinensis View in CoL , M. forcarti View in CoL , M. ginesi View in CoL , M. lipkei View in CoL , M. meansi View in CoL , M. odaelkae View in CoL , M. racenisi View in CoL , M. sucreensis View in CoL , M. turumikiri View in CoL , and M. viloriai View in CoL ). Microthelphusa furcifer View in CoL n. sp. and M. meansi View in CoL occur both in the Potaro-Siparuni Region ( Guyana Shield), although separated by very different altitudes (440 m and 1219 m, respectively). The species of Microthelphusa View in CoL are usually highly endemic, restricted to highlands and with remarkably narrow geographic and altitudinal ranges ( Magalhães 2010; Suárez 2006, table 1); the geographic and altitudinal distribution of Microthelphusa furcifer View in CoL n. sp. and all previously known species of Microthelphusa View in CoL are so far consistent with these patterns ( Table 1 View TABLE 1 ).
We gratefully thanks Rafael Lemaitre (USNM) for a pre-doctoral fellowship to MP and for providing working space and access to the museum collections. We also thanks Karen Reed (USNM) for helping with the collections and bibliographic references and Célio Magalhães (INPA), Ángel Viloria, Carlos Carmona and Héctor Suárez (IVIC) for providing working space and/or access to their respective collections. MP thanks FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo) for providing financial support by way of a doctoral fellowship (2012/ 01334-7). MT thanks CNPq (301806/2010-1) and PETROBRAS (4600224970) for supporting studies on the systematics of decapod crustaceans. This paper greatly benefited from comments from reviewers Célio Magalhães (INPA) and Neil Cumberlidge (Northern Michigan University) and from editor Peter Castro (California State Polytechnic University, Pomona).

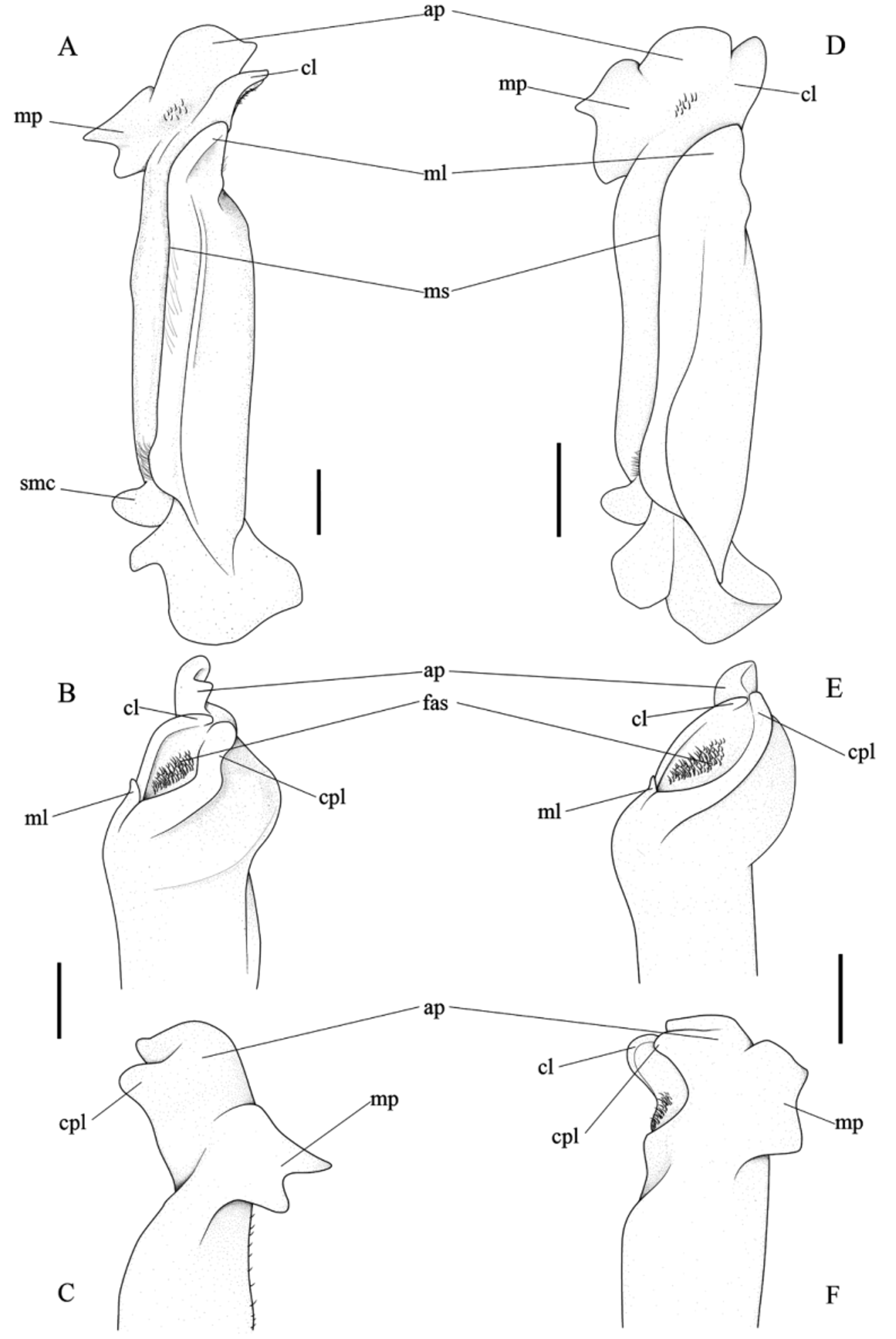
TABLE 1. Geographic and altitudinal distributions for the species of Microthelphusa. Bra, Brazil: Guy, Guyana; Sur, Surinam; Tri, Trinidad; Ven, Venezuela.
| Species | Locality | Altitude (m) |
|---|---|---|
| M. recenisi | Aragua state (Ven) | 1400 – 2000 |
| M. forcarti View in CoL | Mérida state (Ven) | 1603 – 1800 |
| M. sucreensis View in CoL | Monagas and Sucre states (Ven) | 1500 |
| M. turumikiri View in CoL | Sucre state (Ven) | 1500 |
| M. viloriai View in CoL | Trujillo state (Ven) | 1500 |
| M. ginesi View in CoL | Distrito Federal (Ven) | 1400 |
| M. meansi View in CoL | Potaro-Siparuni region (Guy) | 1219 |
| M. lipkei View in CoL | Amazonas state (Bra) | 1133 |
| M. bolivari View in CoL | Bolívar state (Ven) | 1000 |
| M. wymani View in CoL | Paramaribo (Sur) | 880 |
| M. odaelkae View in CoL | Trinidad (Tri) | 600 – 800 |
| M. barinensis View in CoL | Barinas state (Ven) | 530 – 570 |
| M. furcifer View in CoL n. sp. | Potaro-Siparuni region (Guy) | 440 |
| M. rodriguezi View in CoL | Esequive (Guy) | ? |
| M. somanni View in CoL | Amazonas state (Bra) | ? |
| Acknowledgements |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
|
|
Tribe |
Kingsleyini |
|
Genus |