
Spirobolidae Bollman, 1893
|
publication ID |
https://doi.org/ 10.5281/zenodo.5179125 |
|
publication LSID |
lsid:zoobank.org:pub:91F84CDC-6A07-40B6-83F1-35083D171267 |
|
persistent identifier |
https://treatment.plazi.org/id/325C879C-1D74-6335-D1F8-FCCBFB8AF8E6 |
|
treatment provided by |
Felipe |
|
scientific name |
Spirobolidae Bollman, 1893 |
| status |
|
Family Spirobolidae Bollman, 1893 View in CoL
Spirobolinae Bollman, 1893:156 View in CoL . Jeekel, 1971:210 (list). Kevan, 1983:2962.
Spirobolidae View in CoL : Verhoeff, 1893:481; 1931:1680 (key), 1683–1684. Pocock, 1894a:484 (Key); 1894b:388. Cook, 1895:7; 1904:64. Brölemann, 1913:476; 1914:29. Attems, 1926:194, 225. Blake, 1931:16. Chamberlin, 1941:253; 1943a:5; 1947:44; 1949:163; 1951:30; 1953:148. Loomis, 1944:169; 1968a:92; 1968b:391; 1971:53. Hoffman, 1950:31; 1969:233–234; 1980a:76; 1998b:64; 1999:36. Loomis and Hoffman, 1951:51. Causey, 1954:224; 1955ba:87; 1963:78. Johnson, 1954:248 (list). Chamberlin and Hoffman, 1958:159. Keeton, 1960a:28–31. Buckett, 1964:23. Reddell, 1965:164. Stewart, 1969: 384. Filka and Shelley, 1980:16 . Kevan, 1983:2962. Shelley, 1978:52 ; 1988:1642; 2000a:184; 2001a:244; 2002a:98; 2002b:1868. Snider, 1991:179. Wang and Zhang, 1993:845. Watermolen, 1995:2. Wang and Mauriès, 1996:84. Zhang et al, 1997:509. Korsós, 2004:17. Bueno-Villegas et al., 2004:586. Snyder, 2008:18–19.
Euspirobolidae Attems, 1909:25; 1910:89–90.
Type-genus. Spirobolus Brandt, 1833 .
Diagnosis. Small- to large-bodied (primarily moderately large), rounded/cylindrical or transversely ovoid Spirobolidea with the following gonopodal features. Sternum of anterior gonopods relatively narrow and arched mediad, with or without medial lobe protruding ventrad between coxal endites. Latter large and prominent, overlying and closely appressed to sternum, either separated by medial sternal lobe or (sub)contiguous. Telopodites moderately long, curving mediad, either fused to coxa with only faint suture line or separate from and articulating with latter; apex usually slightly prolonged and variably configured, either linear, rounded, or subacuminate, uncinate or not. Posterior gonopod sternum usually absent, a vestigial remnant in one tribe. Coxa short; apodeme spatulate, varying from short and oriented dorsoventrad to long and extending caudad inside body up to 10 th ring. Telopodite with variable prefemoral process, either an inconspicuous, nubbin-like enlargement on telopodital stem or a variably long, lobate, conspicuous projection subequal in length to acropodite. Latter long and prominent, overall appearance varying from thick, upright, and post-like to a thin, laminate, endomere-like hood leaning strongly mediad and overhanging and enveloping prefemoral process to varying degrees, caudal and/ or anterior surfaces extending submediad for short but varying lengths, margins thickened and lobate with or without additional projections, or thin, laminate, sublinear and with or without indentations, marginal lobes, and spiniform projections.
Components. Two subfamilies: Floridobolinae Keeton, 1959 , and Spirobolinae Bollman, 1893 .
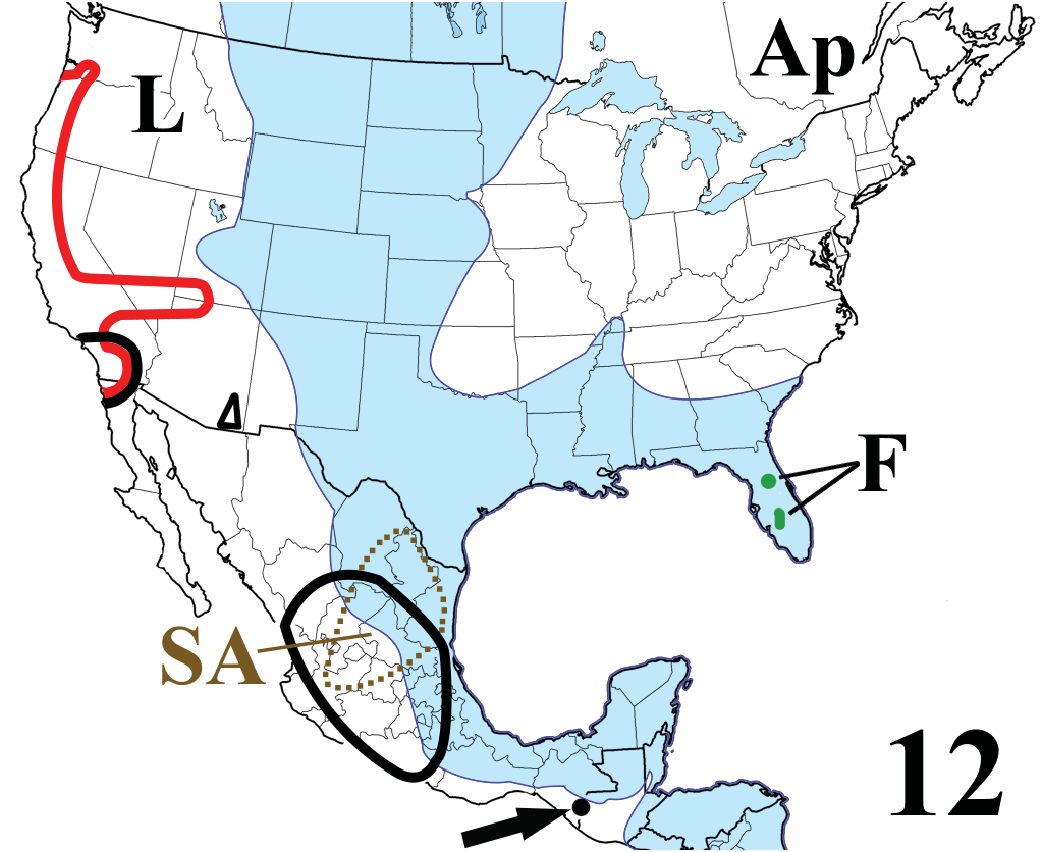
Distribution ( Fig. 11 View Figure 11 ). Occupying seven variably disjunct areas, six in North and Central America and one in eastern Asia. In eastern North America, Spirobolidae range from southern Québec, Canada, and Maine and the Upper Michigan Peninsula, USA, to the south Florida Keys and the Gulf Coast to the latitude of Nueces Bay/Corpus Christi, Texas; east-west, they extend from the Atlantic Ocean from Maine through Florida to the Plains of eastern Nebraska and central Texas, with an apparently allopatric population some 80 km (50 mi) to the northwest in southwestern Minnesota. In western North America, Spirobolidae occur continuously between the Pacific Ocean and the crest of the Sierra Nevada and Cascade Mountains from north of the Columbia River in Klickitat and Sakamania cos., Washington, USA, to northern Baja California Norte, Mexico, with a dactyliform eastward expansion from the southern Sierra Nevada through the Mohave and Great Basin deserts, Nevada, to southwestern Utah and northwestern Arizona. The fourth US area is a small, isolated, subtriangular region in the Pinaleno, Dragoon, and Chiricahua Mountains of southeastern Arizona. In “mainland” Mexico, Spirobolidae occupy a broad, central area that extends, north-south, from northern Nuevo León and central Durango to Guerrero, with an isolated point locality on Volcán Tajumulco, San Marcos Dept., Guatemala ( Chamberlin and Hoffman 1958; Keeton 1960a, 1966; Loomis 1968a; Hoffman 1980a, 1999; Shelley 1988 , 2001a, 2002a, c, 2010; Shelley and Hoffman 1995; Shelley and Bauer 1997; Shelley and Stevens 2003; Bueno-Villegas et al. 2004; Shelley et al. 2006; Shelley and Golovatch 2011; plus unpublished records cited in the Appendix). The east Asian area covers Taiwan and eastern China from the East China Sea to Sichuan Province (Prov.) and from north of Beijing to Guizhou Prov. ( Wang and Mauriès 1996, Korsós 2004), with a disjunct fossil record in southern Mongolia. Distances between presently occupied areas are listed in Table 3.
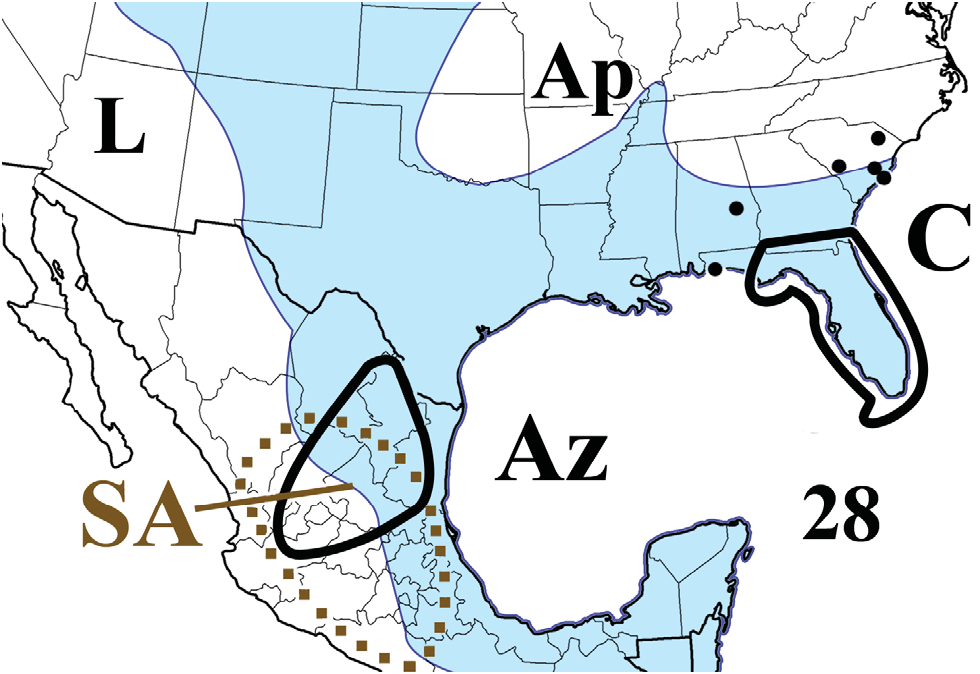
Origin. Represented by Floridobolini , Spirobolidae arose in northern “proto-Mexico” (“northern Mexican Highlands” [ Keeton 1960a]), after Euramerica merged with the “proto-South America” part of “Gondwana I” around 306 mya. Their geographically restricted distributions suggest that Allopocockiidae , Atopetholidae , Hoffmanobolidae , Messicobolidae , and Typhlobolellidae also originated there after this geologically dated event. We submit that this source area is plausibly the mountainous region of northern Mexico where Floridobolinae (Tylobolini) and Spirobolinae (Aztecolini) overlap ( Fig. 12 View Figure 12 , 28 View Figure 28 , 32 View Figure 32 ), which includes all of Aguascalientes and parts of Coahuila, Guanajuato, Jalisco, Nuevo León, San Luis Potosi, Tamaulipas, and Zacatecas states. Allowing a modicum of time for Floridobolini to evolve from ancestral spirobolidean stock, we arbitrarily estimate the age of the family and Floridobolinae / ini at <303 my.
Remarks. Though specialized in some ways, spirobolid posterior gonopod telopodites are simpler than those in other spirobolidean families, involving only a prefemoral process and a dominant acropodite, both varying in size, position, proportions, and configuration. Noting the family’s discontinuous distribution and restriction to temperate latitudes in a primarily tropical (sub)order, Keeton (1960a) concluded that it held relictual status and was unable to withstand competition from more advanced tropical families. This, however, is not the case; Spirobolidae and the families in the previous paragraph arose in the general northern Mexican source area where elements of each remain today.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Spirobolidae Bollman, 1893
| Shelley, Rowland M. & Floyd, Samuel D. 2014 |
Spirobolinae
| Kevan, D. K. 1983: 2962 |
| Jeekel, C. A. W. 1971: 210 |
| Bollman, C. H. 1893: 156 |
Spirobolidae
| Snyder, B. A. 2008: 18 |
| Korsos, Z. 2004: 17 |
| Bueno-Villegas, J. & P. Sierwald & J. E. Bond 2004: 586 |
| Shelley, R. M. 2002: 98 |
| Shelley, R. M. 2002: 1868 |
| Shelley, R. M. 2001: 244 |
| Shelley, R. M. 2000: 184 |
| Hoffman, R. L. 1999: 36 |
| Hoffman, R. L. 1998: 64 |
| Zhang, C. & D. Wang & F. Zhang 1997: 509 |
| Wang, D. & J. - P. Mauries 1996: 84 |
| Watermolen, D. J. 1995: 2 |
| Wang, D. & C. Zhang 1993: 845 |
| Snider, R. M. 1991: 179 |
| Shelley, R. M. 1988: 1642 |
| Kevan, D. K. 1983: 2962 |
| Hoffman, R. L. 1980: 76 |
| Filka, M. E. & R. M. Shelley 1980: 16 |
| Shelley, R. M. 1978: 52 |
| Loomis, H. F. 1971: 53 |
| Hoffman, R. L. 1969: 233 |
| Stewart, T. C. 1969: 384 |
| Loomis, H. F. 1968: 92 |
| Loomis, H. F. 1968: 391 |
| Reddell, J. R. 1965: 164 |
| Buckett, J. S. 1964: 23 |
| Keeton, W. T. 1960: 28 |
| Chamberlin, R. V. & R. L. Hoffman 1958: 159 |
| Causey 1954: 224 |
| Johnson, B. M. 1954: 248 |
| Chamberlin, R. V. 1951: 30 |
| Loomis, H. F. & R. L. Hoffman 1951: 51 |
| Hoffman, R. L. 1950: 31 |
| Chamberlin, R. V. 1949: 163 |
| Chamberlin, R. V. 1947: 44 |
| Loomis, H. F. 1944: 169 |
| Chamberlin, R. V. 1943: 5 |
| Chamberlin, R. V. 1941: 253 |
| Verhoeff, K. W. 1931: 1680 |
| Blake, C. H. 1931: 16 |
| Attems, C. 1926: 194 |
| Brolemann, H. W. 1914: 29 |
| Brolemann, H. W. 1913: 476 |
| Cook, O. F. 1904: 64 |
| Cook, O. F. 1895: 7 |
| Pocock, R. I. 1894: 484 |
| Verhoeff, K. W. 1893: 481 |