
Boulengerula spawlsi, Wilkinson, Mark, Malonza, Patrick K., Campbell, Patrick & Loader, Simon P., 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4286.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:89438641-D8A5-42D1-88A1-0C495E139E3B |
|
DOI |
https://doi.org/10.5281/zenodo.6032782 |
|
persistent identifier |
https://treatment.plazi.org/id/934C4926-5EB0-467E-83AB-E81E96A9358E |
|
taxon LSID |
lsid:zoobank.org:act:934C4926-5EB0-467E-83AB-E81E96A9358E |
|
treatment provided by |
Plazi |
|
scientific name |
Boulengerula spawlsi |
| status |
sp. nov. |
Boulengerula spawlsi sp. nov.
urn:lsid:zoobank.org:pub:FCA5F156-422B-4E07-ACF1-189F6BFA873F ( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 , Table 1)
Boulengerula cf. denhardti Nieden, 1912 : Loader et al. (2011: 6, figure 1)
Boulengerula denhardti Nieden, 1912 : Measey et al. (2012: 187 –191, figure 1)
Holotype. NMK A5596/2 (MW 7938), an adult female collected by the authors in Ngaia Forest Reserve , Nyambene Hills, Meru County, Kenya, N 0° 22 ' 47.3 '' E 38° 0 ' 47.5 '', 1419 m asl, 23rd January 2013. GoogleMaps
Paratypes (n = 8). BMNH 2005.2264 (formerly NMK A5596/1, MW 7937) same data as holotype except N 0° 22 ' 44.8 '' E 38° 0 ' 45.9 '', 1406 m asl GoogleMaps ; BMNH 2005.2265 (formerly NMK A4954/4 / PKM00871), A4954/1 (PKM00847), A4954/2 (PKM00849), A4954/3 (PKM00851), A4954/5 (PKM00873), A4954/6 (PKM00875) collected near the type locality (N 0° 23 ' 19.3 '' E 38° 0 1 ' 41.6 '', 1334 m asl) by PKM and Vincent Muchai, 22–23 May 2008; A/4824 (PKM00622) collected near the type locality (N 0° 19.063 ' E 38° 0 0.06 ', 1300 m asl) by Stephen Spawls 13th February 2007.
Diagnosis. A Boulengerula that differs from all congeners in having the anterior part of the tentacular canal covered by maxillopalatine. It differs further from B. boulengeri Tornier, 1896 and B. denhardti in having inner mandibular teeth, from B. changamwensis Loveridge, 1932 and B. fisheri Nussbaum and Hinkel, 1994 in having a mainly darker blue or purple, rather than pink, body colour, and from all other Boulengerula ( B. taitanus, Loveridge, 1935 , B. niedeni Müller, Measey, Loader & Malonza, 2005 , B. uluguruensis Barbour & Loveridge, 1928 ) in having more (> 150) annuli.
Description of the holotype. Some morphometric and meristic data are in Table 1. Good condition except for some wrinkling and creasing of the skin, a 5 mm midventral incision c. 90 mm behind ST, a 7 mm midventral incision c. 43 mm anterior of TT.
Body subcylindrical, slightly dorsoventrally flattened, fairly uniform, narrowing noticeably only in the region of the vent and terminal shield; L/W c. 66. In dorsal view, head more U- than V-shaped; sides of head fairly straight and converging substantially from the back of the head to about halfway between TAs and nares, and more strongly anteriorly to a bluntly rounded ST. In lateral view, head wedge-shaped; top of and bottom of head fairly straight and converging strongly up to level of nares and then more curved and strongly converging to narrow rounded tip; upper lip very slightly concave, slightly downturned close to AM; ridge bearing vomeropalatine teeth visible close to CM; lower jaw about one half and one fifth the height of upper jaw at levels of CM and TA respectively. ST much blunter in dorsal than in lateral view. In ventral view, snout projects strongly beyond recessed mouth, anterior margins of upper and lower jaws much more bluntly rounded than ST, sides of upper jaw visible as far back as CM. Eyes not visible. Subcircular TAs very close to lip, slightly elevated, on distinct subcircular TPs, that are just visible dorsally and more so ventrally, much closer to nares than to CMs, below (touching) imaginary lines between nares and CMs, slightly (one TP diameter) posterior to level of AM. Ovate nares small, approximately the same size as TAs, dorsolateral, closer to ST than to level of AM, about as far from bottom as from top of snout and half as far from ST in lateral view, barely visible from below a little more so from above.
Teeth pointed, gently recurved, last few elements of outer mandibular series much smaller. OMs and PMs monocuspid, the former larger; VPs smaller, bicuspid, vomerine series broadly rounded anteromedially, palatine series extending posteriorly much further than premaxillary-maxillary series, cuspidity of paired IM teeth unclear. Distance between vomeropalatine and premaxillary-maxillary series anteriorly much less (approximately half) AM–ST in ventral view; upper teeth series extending posteriorly to about the level of posterior margin of choana. Palate not strongly arched transversely or longitudinally. Choanal apertures subcircular, anterior margins far behind level of TAs, separated from each other by about twice width of single choana. Tongue smooth, somewhat crescent shaped with concave margin posterior, rounded and unattached anteriorly.
Nuchal region scarcely wider than adjacent body and head. Two nuchal collars clearly marked by three nuchal grooves; NG1 and NG2 completely encircling body, NG3 widely incomplete (or at least hard to discern) on the venter. Substantial TG on C2, visible laterally. All grooves in nuchal region bow forward medially on dorsum. NG1 bowing slightly forward ventromedially. A small midventral crease extends from close to the mentum to just past the ventral TG on C1. AGs mostly incomplete dorsally except first 12 and the first 14 of the last 19, complete or narrowly incomplete ventrally; last seven or eight annuli shorter than the others. Last three AGs increasingly difficult to discern, interrupted ventrally by the vent and disc; last approximately level with vent. Body ends in a short terminal shield approximately twice the length of the last annulus (viewed laterally), a little shorter than (3/4 the length of) midbody annuli. Distinct terminal keel on dorsal surface of terminal shield. Body terminus bluntly rounded in dorsal view, narrowing only in the vent region. Rounded in lateral view, ventral surface somewhat flattened. Vent circular, towards the back of an eliptical disc that is longer than wide (1.8 x 1.3 mm), perhaps five main denticulations anterior and four posterior, but poorly indicated and with irregular subdivisions, the interdenticular creases reaching to margins of disc and thus longer anteriorly; no papillae. Three vertebrae in the nuchal region, no post-cloacal vertebrae.
TABLE]. Morphometric (in mm) anđ meristic đata for Boulengerula spawlsi sp. nov. Entries in parentheses are from Measey et al. (2012). * đenotes holotype, j = juvenile. Circumference measuređ at miđbođy.
In preservative a slightly bluish olive grey, darker and more blue narrow (2.7 mm wide) dorsal band not extending laterally onto bulges produced by the underlying superficial external oblique trunk muscle, especially dark along AGs and, anteriorly along dorsal midline. Somewhat paler ventrally, with some much paler patches on flanks especially near the head and terminus. AGs marked by a broad dark band with a medial row of whitish glands visible with magnification. Tip of snout dorsally and entire ventral surface of head pale and unpigmented as far posterior as NG2. TPs encircled by slightly darker narrow ring. Ventral surface of terminal area pale, disk cream.
In life ( Fig. 2 View FIGURE 2 A–B the holotype was a slightly purplish blue dorsally, slightly paler on the flanks and below with a pinkish head and throat and a pale snout tip. AGs were whitish with this pale colour extending slightly laterally and ventrally on the last few annuli and also surrounding the slightly darker disc.
Variation and additional information from paratypes. Variation in some meristics and morphometrics is summarised in Table 1. The paratypes agree in most aspects with the holotype. All have a pair of inner mandibular teeth. The most conspicuous variation is in the extent of the pale colour on the throat region which extends further posteriorly onto the collars in some specimens. Most paratypes are a little more dehydrated than the holotype, resulting in some shrinkage and the artefactual appearance (in X-radiographs) of vertebrae behind the level of the vent in some, as has been reported in other species ( Nussbaum 1988). One paratype (NMK A4824) has the jaws broken, facilitating determination that the IMs are bicuspid.
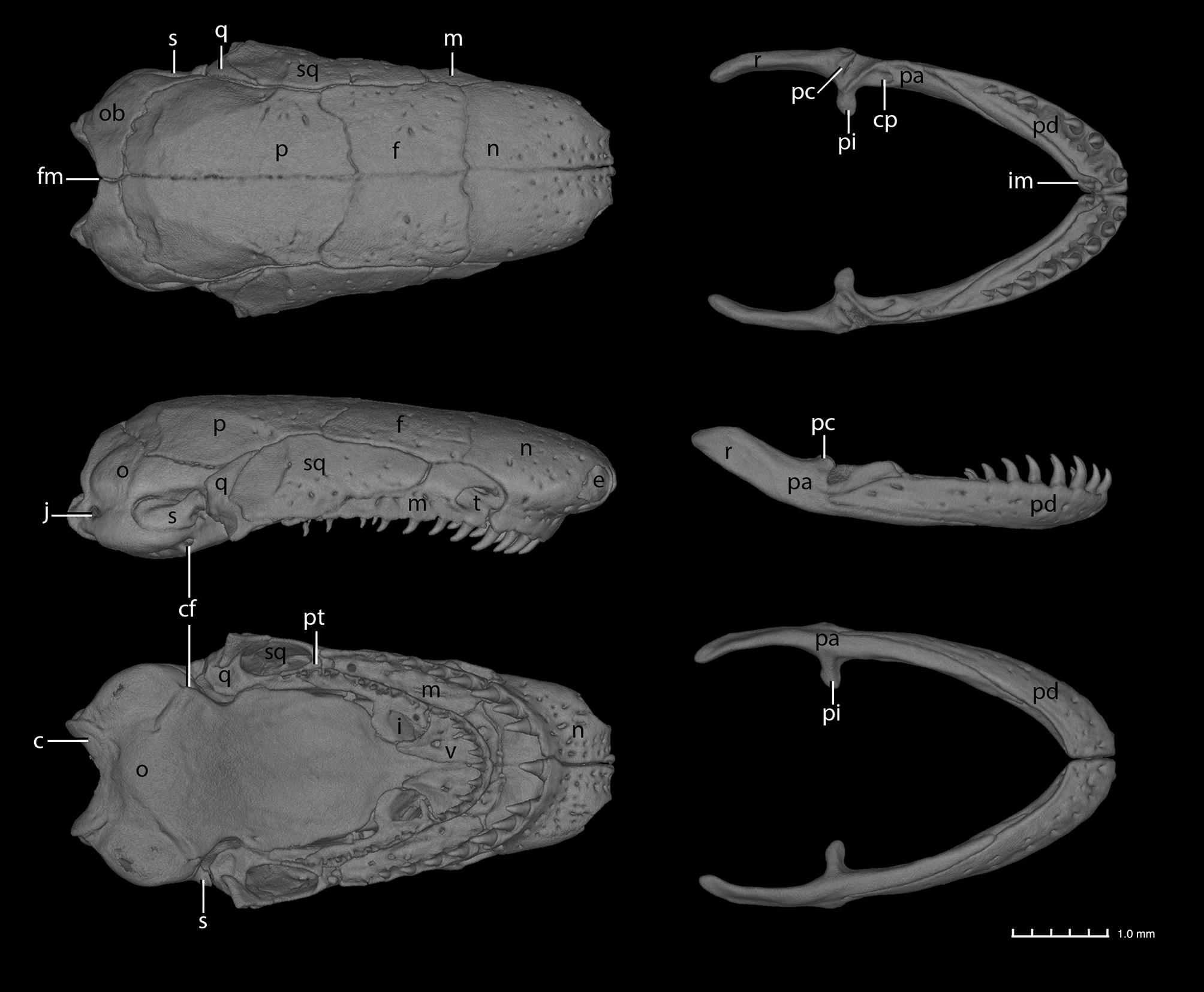
CT scans of BMNH 2005.2264 ( Fig. 3 View FIGURE 3 ) and two other paratypes (NMK A4954/5 and A4954/6, data not shown) reveal details of the osteology of the new species. The general composition, shape and arrangement of bones is as in other Boulengerula ( De Villiers 1938; Taylor 1969, MW pers. obs.) with the exception, seemingly correlated with the relatively anterior placement of the TA, that the maxillopalatine forms a complete or partial bony covering of the anterior part of the tentacular groove. Other notable features are (1) comparatively weak diastemata between the vomerine and palatine teeth series on each side, (2) PM series that are substantially shorter than (do not extend as far back as) the VP series, (3) a pair of inner mandibular teeth, (4) no exposure of the mesethmoid dorsally, (5) separate (paired, left and right) foramina in the sphenethmoid for the passage of the dorsal roots of the olfactory nerve, and (6) imperforate stapes. Three of these (1, 5 and 6) are additional substantial differences between the three scanned specimens of the new species and the holotype of B. denhardti (MW pers. obs.). In all cases, tooth counts made directly from specimens agreed very closely with those made from CT scans (data not shown).
Remarks. The holotype was dug from soil in a hollowed out base of the trunk of a large fallen tree ( Fig. 2 View FIGURE 2 C) and NMK A5596/1 was collected in the soil beneath a rotting log, these two specimens collected in approximately 12 person hours of digging. NMK A4824 was found under a stone. Large decomposing logs are not very abundant in the forest and not all of them harbour caecilians. The forest is used by the local community and log collecting for firewood may negatively impact the extent of suitable microhabitat for Boulengerula spawlsi . This species does not appear to be as abundant (or at least not as readily found) as some of its congeners. Some other Kenyan Boulengerula , for example B. taitanus and B. niedeni , are more readily found in farmland than in forest ( Malonza & Measey, 2005; Malonza et al., 2010) but whether B. spawlsi occurs outside of forest remains to be determined. The presence of B. spawlsi in a protected areas implies some protection for the species, but the extent of its distribution in the forests of the Nyambene Hills (e.g. in Igembe, and Kageta forest blocks that are at higher elevations than Ngaya) and in farmlands outside the forest needs to be better understood to usefully inform its conservation status.
Etymology. The species is named in honour of Stephen Spawls in recognition of his substantial contributions to African herpetology, including his discovery of this species of caecilian.
Suggested English name. Spawls’ boolee.


| NMK |
National Museums of Kenya |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Boulengerula spawlsi
| Wilkinson, Mark, Malonza, Patrick K., Campbell, Patrick & Loader, Simon P. 2017 |
Boulengerula denhardti
| Measey 2012: 187 |
Boulengerula cf. denhardti
| Loader 2011: 6 |