
Sinobatis
|
publication ID |
https://doi.org/ 10.5281/zenodo.180189 |
|
DOI |
https://doi.org/10.5281/zenodo.5672360 |
|
persistent identifier |
https://treatment.plazi.org/id/3109F354-FFFF-1730-99F5-FE3C0671FE4A |
|
treatment provided by |
Plazi |
|
scientific name |
Sinobatis |
| status |
|
Genus Sinobatis View in CoL View at ENA
Sinobatis Hulley 1973:153 View in CoL . Fem. Anacanthobatis borneensis Chan 1965 View in CoL . Type by original designation.
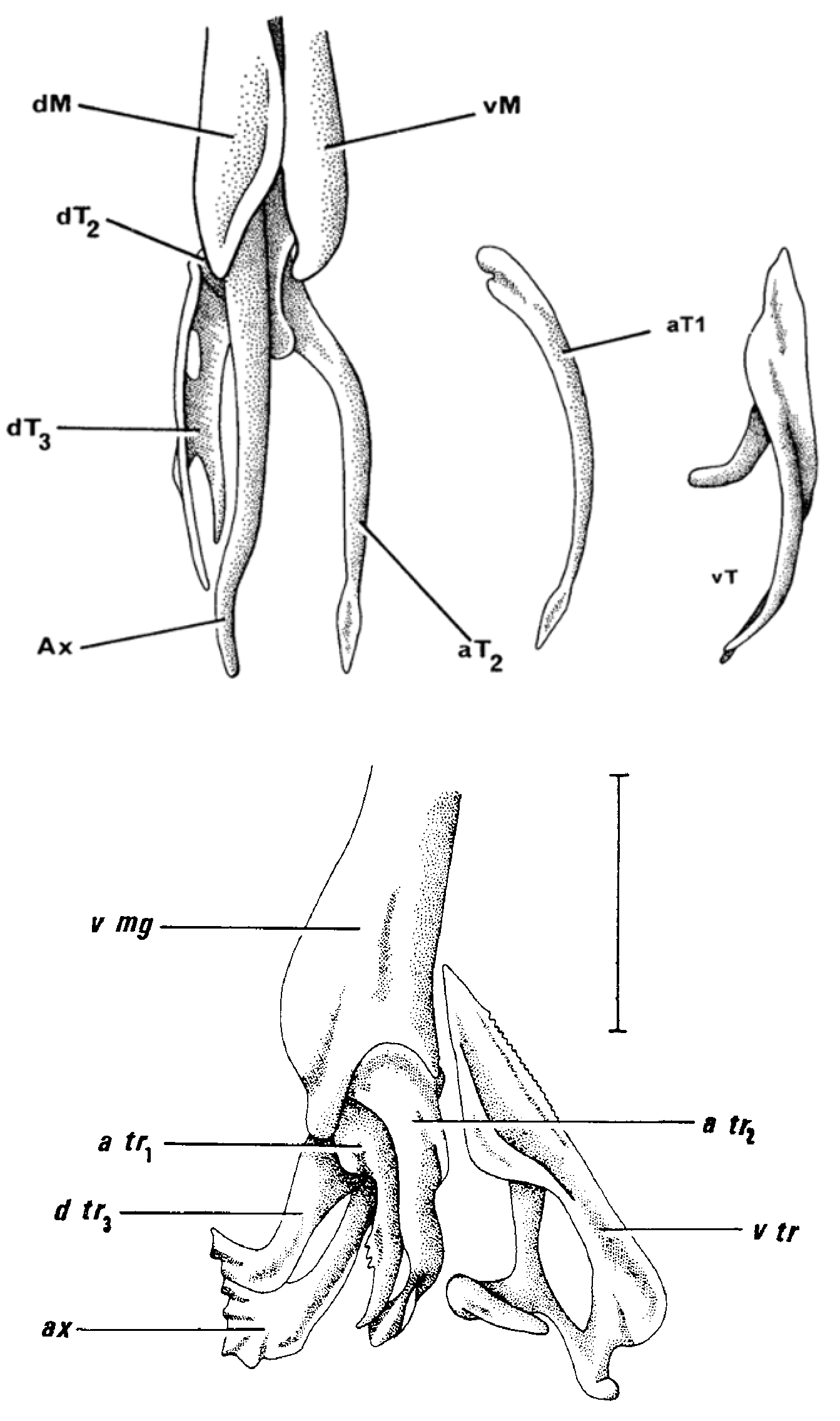
Definition. Claspers small to medium (postcloacal length about 17–21% TL when mature); lacking spur, palp and pseudosiphon; rhipidion present; shield narrow laterally, with an expanded medial arm that wraps over ventral surface and fuses to axil cartilage; sentinel and spike simple with pungent, slightly flattened, recurved tips; sentinel capable of strong anteromedial rotation (almost forming 180° angle with spike when clasper everted); two dorsal terminal cartilages but dorsal terminal 1 absent; distal projection of dorsal terminal 3 not forming an obvious spur; axial tip simple, not spatulate or with a corrugated margin; accessory terminal 1 simple, apex weakly or strongly blade like, not spoon-shaped; accessory terminal 2 simple, slender, forming a spiral projection, attachment process poorly developed; monospondylous vertebrae mainly exceeding 25.
Remarks. Sinobatis is elevated to generic level on the basis of the conservative intragroup clasper morphology of its member species, and its strong differentiation from Hulley’s (1973) other subgenera of Anacanthobatis . The adult claspers of two of the new species, for which adult males were available, conform very closely to Hulley’s figures and definition of Sinobatis based on S. borneensis ( Fig. 1 View FIGURE 1 ). These species form a natural group based on their clasper morphology. The clasper of the type species of the genus Anacanthobatis , A. marmoratus (Von Bonde & Swart, 1923) , differs markedly from these 3 species ( Fig. 2 View FIGURE 2 , see also Hulley, 1972). Anacanthobatis has a palp (absent in Sinobatis ), prominent shield with a well-developed eperon (poorly developed without an eperon), spatulate axial tip (simply pointed), 3 dorsal terminal cartilages (only 2 with dorsal terminal 1 absent), spur present (absent), and ventral terminal cartilage with serrated outer lateral margin and windowed inner lateral margin (no serrations and without fenestration). Hulley (1972) initially described and figured the accessory terminal 2 of Anacanthobatis as spoon-shaped distally but later relabelled this structure as the accessory terminal 1 ( Hulley, 1973). In Sinobatis , both of these cartilages are slender and pointed distally.
We recommend that Hulley’s two other subgenera, Springeria and Schroederobatis , be elevated to generic level as their clasper morphology also differs markedly from Anacanthobatis and Sinobatis . Springeria (based on Hulley, 1973) differs from Sinobatis in possessing a spur and palp (otherwise absent), the shield is well developed (poorly developed in Sinobatis but lacking an eperon in both taxa), four dorsal terminal cartilages (rather than 2), ventral terminal cartilage with a small medial process (process greatly extended, wrapping around clasper), and the accessory terminal 2 with a much better developed attachment process. Schroederobatis (based on Hulley, 1973) differs from Sinobatis in having a pseudosiphon, but lacks a shield and ventral terminal cartilage. Also, the accessory terminal cartilages are reduced in length in Schroederobatis , and the dorsal terminal 1 is well developed (rather than absent). For more details see Hulley (1972, 73).
Other taxonomic characters appear to be important for characterising these groups. For example, Séret (1986) has shown that there are major differences in the chondrocranium between members of the subgroups. Similarly, species of Sinobatis have a relatively short and broad nasal curtain with rather small lateral lobes compared to the long, narrow, large-lobed curtain possessed by Springeria longirostris . More work is required to provide comprehensive definitions of the genera and assign presently unresolved taxa such as Anacanthobatis ori (Wallace, 1967) which, based on its chondrocranium, is unlikely to be congeneric with A. marmoratus .
A B Species. Sinobatis includes S. borneensis (Chan) (South China Sea and Taiwan), S. bulbicauda sp. nov. (eastern Indonesia and northwestern Australia, SE Indian Ocean), and S. filicauda sp. nov. (northeastern Australia, SE Pacific Ocean) based on observed adult clasper morphology, and provisionally includes S. melanosoma (East and South China Seas and Taiwan) and S. caerulea sp. nov. (northwestern Australia, SE Indian Ocean). The 317 mm TL, mature male holotype of Sinobatis borneensis (BMNH 1965.1.29.1) was collected in the South China Sea, north of Kuching, Borneo. Compared to other members of the genus, S. borneensis appears to be relatively small. An adolescent male examined from the nearby Philippines (MNHN 1997– 3321) had well-developed claspers and developing alar thorns at 253 mm TL. Chu et al.’s (1981) figure of the late adolescent male holotype of Springeria nahaiensis , also from the South China Sea, displays developing claspers and alar thorns at 295 mm TL, and is probably a juvenile of S. borneensis . Another larger male Philippine anacanthobatid (CSIRO H 4125–01) had undeveloped claspers at 382 mm TL. It is referable to Springeria melanosoma Chan from the South China Sea based on its dark dorsal and ventral coloration and black areas around the spiracles and orbits, body shape, and relatively well-developed caudal fin. Four large Sinobatis (455–594 mm TL, males still immature at 528 mm TL) collected from the Okinawa Trough and identified by Ishihara (1984) as S. borneensis are clearly too large for that species. Figures of a large adult male (MTUF 25001, 551 mm TL), and a larger female (MTUF 25004, 594 mm TL), are closer to S. melanosoma . Ishihara’s paper does not provide an expanded image of the adult claspers but these appear to be of the Sinobatis type with the spike and sentinel forming an angle of about 180° to each other when the clasper glans is everted. Further work is needed to resolve the alpha taxonomy of western North Pacific anacanthobatids.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Sinobatis
| Last, Peter R. & Séret, Bernard 2008 |
Sinobatis
| Hulley 1973: 153 |