
Bathyconchoecia darcythompsoni ( Scott, 1909 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3254.1.2 |
|
persistent identifier |
https://treatment.plazi.org/id/3034D544-FFD4-C64D-FF52-FE6B66ACFAA0 |
|
treatment provided by |
Felipe |
|
scientific name |
Bathyconchoecia darcythompsoni ( Scott, 1909 ) |
| status |
|
Redescription of Bathyconchoecia darcythompsoni ( Scott, 1909) View in CoL
( Figs 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Synonymy
1909 Euconchoecia d’arcy-thompsoni Scott: 128–129, Pl. IV, figs 1–12.
1968 Bathyconchoecia darcythompsoni ─ Deevey : 540.
1969a Bathyconchoecia darcythompsoni ─ Poulsen : 142–3, fig. 6.
1972 Bathyconchoecia darcythompsoni ─ Poulsen : 450–451, fig. 3.
1993 Bathyconchoecia darcythompsoni ─ Angel : 86, fig. 29.
The carapace lengths of all the adults and juveniles instars collected are summarised in Table 1.
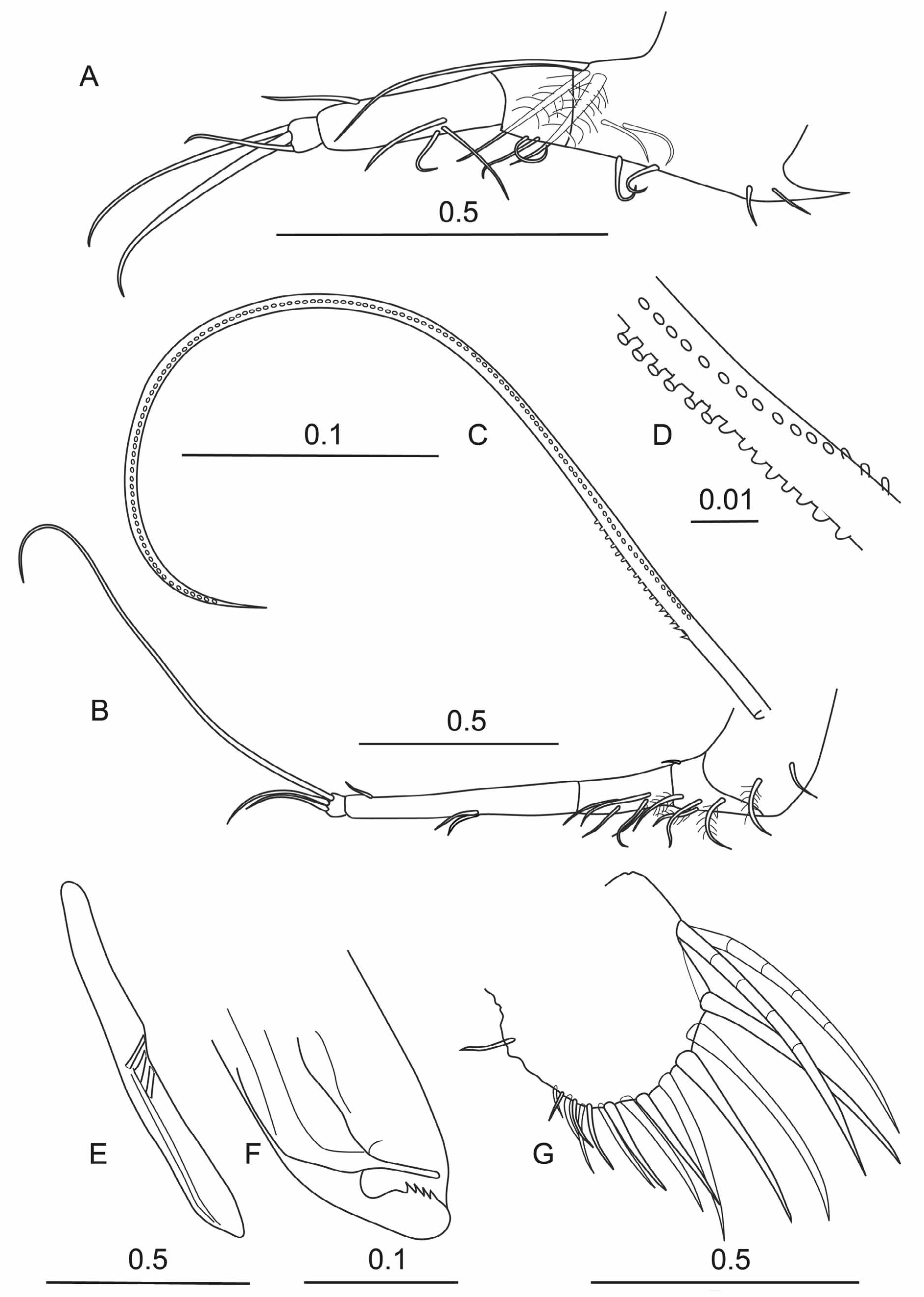
Male ( Fig. 5 View FIGURE 5 ). The carapace is long relative to its height and breadth. The height is about 44 % CL and the breadth is 38 % CL. The lateral outline ( Fig. 5B View FIGURE 5 ) of the carapace is very much similar to Scott’s original drawings (Plate IV, figs 1 and 2). The surface of the carapace is sculpted with a faint scaly pattern with fine pits. In the anterior region beneath the incisure, the pits become aligned to form a series of striae that initially intercept the ventral margin, but then run parallel to the ventral margin. However, this pattern of striation is not evident in all specimens. The rostrum is long, about 13.2 % CL when measured from the anterior end of the hinge between the two carapace valves and the tip of the rostrum. A feature unique to males of this species is a 0.24 mm deep sulcus that divides the rostrum in two (as illustrated by Scott).
The incisure is deep and broad (16.6 % CL from the tip of the rostrum to the inner margin). The dorsal margin of the carapace is almost straight and there is only slight development of shoulder vaults. The ventral margin is almost symmetrically curved with the maximum carapace height at midlength and it curves smoothly into the posterior margin. The posterior dorsal corner of the left valve is a rounded right angle, but on the right valve there is a small point. The carapace glands are large, and occupy almost a third of the carapace height below the posterior dorsal corner. Each gland opens within a curved slot, which is situated in an infold 25 % of the carapace height below the posterior dorsal corner. These symmetrical glands are probably homologous to the asymmetrical carapace glands in the Conchoeciinae .
Frontal organ. There is no frontal organ.
First antenna ( Fig. 5D View FIGURE 5 ). The first antenna is indistinctly 6-segmented (but note Poulsen’s (1969b) interpretation that there are seven segments). The limb arches ventrally and the basal three segments are bare. On the ventral side of the fourth segment is the large ventral pad on which are mounted about 8 rows each of about 30 fine setae (26.9 % CL). The fifth segment carries a plumose dorsal seta. The terminal segment carries four setae one of which is very long (~40 % CL). Scott (1909 pl. 4, fig. 5) shows the two longest setae as being of similar length, but it is likely that in his specimen the longest seta was broken—as it was in this specimen.
Second antenna ( Fig. 5E, F, G View FIGURE 5 ). The protopodite of the second antenna is drop-shaped (39.2%CL). The first exopodite segment is slightly curved and is half the length of the protopodite (19.5 % CL). No terminal seta was seen on the first segment in the male. The combined length of the eight other exopodite segments that carry the swimming setae is 7.6 % CL. The first endopodite segment is elongate and rectangular; it is nearly a third the length of the protopodite (12.3 % CL), and its two terminal setae are relatively long. The a-seta is 5.8 % CL and carries fine spinules along its whole length. The b seta is even longer (7.3 % CL) and has long setules near its base which abruptly become shorter distally; the distal third of this seta is bare. The second endopodite segment is about half the length of the first and carries two very long terminal setae (41.2 and>58.8 % CL respectively). The third segment forms the hook appendage which has a straight hasp on the end of which are inserted three unequal setae (>19.5, 12.3 and 6.7 % CL). On the right limb ( Fig. 5F View FIGURE 5 ) the hook beyond the hasp is bent back about 120° and the distal arm curves to the ridged rounded ending, forming a structure similar to a shepherd’s crook or crummock. On the left limb, the distal end piece is a much weaker structure ( Fig. 5G View FIGURE 5 ), but is similar in shape.
Mandible ( Fig. 6A, B, C View FIGURE 6 ). The structure of the mandible is not sexually dimorphic. It is very similar to Scott’s (1909) illustration. The first endopodite segment has a long bare terminal seta that extends to the end of the limb, which is much longer than the homologous setae in Conchoeciinae . On the inner surface of the segment are three subequal setae. On the dorsal edge of the second segment there are three unequal terminal setae, the longest of which is spinous along much of its length. On the ventral surface of the segment is a single medial seta (c.f. two subequal terminal setae in Conchoeciinae ). The terminal segment carries seven terminal setae, the longest of which is as long as the three segments of the endopodite (in the male this seta was broken but in females the longest seta is 21.7 % CL).
The structure of the toothed edge of the basale ( Fig. 6B View FIGURE 6 ) is typical for the genus, and is markedly different from that of Bathyconchoecia paulula , as illustrated, albeit rather indistinctly, by Deevey (1968, fig. 2b). In B. paulula the structure is similar to that of most Conchoeciinae with two spine teeth offset from six cutting teeth, the first of which is also slightly offset and there is an inner tooth, which overlaps two or three of the cutting teeth. In B. darcythompsoni the two small spine teeth are on a level with the first of the cutting teeth, but are well offset from the five cutting teeth ( Fig. 2 B View FIGURE 2 ). The outermost of these cutting teeth is separated from the other four by a sulcus and is possibly homologous with the inner tooth of Conchoeciinae . The other tooth lists are also strikingly different. The outer list consists of two large teeth and five smaller teeth and an inner list that carries rather several blunt teeth arranged in two ranks. The masticatory pad is hemispherical and is covered with short stubby grinding spines and fringed by short, fine spines. The coxale has one latral and three outer setae close to the toothed edge and a long subterminal seta that is finely plumose (not illustrated). On the dorsal terminal margin there are three long plumose setae but no process; in Conchoeciinae there is usually a process that carries a plumose seta, which has been interpreted as being a vestigial endopodite ( Boxshall 1998).
Maxilla ( Fig. 6D View FIGURE 6 ). Scott’s (1909) figure (pl. IV, 8) does not show the segmentation clearly. There are the two usual segments subtended at the base by a plumose seta. On the anterior face of the first segment there is a proximal group of four setae and a single distal seta, all of which are plumose. On the posterior face there is a row of four plumose setae. The distal end of the outer face is bare. The terminal segment has two terminal claw setae subtended by a pair of slim setae on both sides.
Fifth limb ( Fig. 5H View FIGURE 5 ). There are substantial differences between the limb illustrated in Fig. 5H View FIGURE 5 and Scott’s (1909) plate IV, 9. Firstly, assuming that the long dorsal seta is the residual endopodite, and so is inserted on the basale, there are three exopodite segments (compared with two in Conchoeciinae ). The long seta is also subtended by three plumose setae. The first limb segment carries a fairly short dorsal seta and a pair of medial seta ventrally. The long slim second segment also has a medial pair of ventral setae, a medial lateral seta and a dorsal seta at twothirds length. The third segment carries the usual three terminal setae, the longest of which, the central seta, is 10.7 % CL.
Sixth limb ( Fig. 5I View FIGURE 5 ). Scott’s drawing of the sixth limb is almost identical to Fig. 5I View FIGURE 5 . The setation of the basale is identical, with the dorsal seta being quite short and extending only to the middle of the first segment. However, Scott’s illustration shows a single additional seta with the two pairs of ventral setae on the first segment. The second segment has a pair of setae on the ventral surface at one third length, and a dorsal seta at two-thirds length. The third terminal segment has three terminal setae. The dorsal seta is exceptionally long (26.4 % CL), as is a characteristic of the new genus, but is not armed as in S. arabica .
Caudal furca. There are eight pairs of hook setae, but all were broken in the dissected male specimen, but they are likely to be similar to those illustrated for the female ( Fig. 7I View FIGURE 7 ).
Copulatory appendage ( Fig. 5J View FIGURE 5 ). The appendage is a single structure inserted on the left side of the caudal furca. It is slim, curved and long (17.5 % CL) as illustrated by Scott (1909, fig. 12) with a rounded end. Scott’s (1909) illustration does not show the five oblique muscles in the base. The terminal stylet is rather obscure.
Female. Carapace ( Fig. 7A View FIGURE 7 ). The female carapace is on average slightly shorter than the male’s (see Table 1) but is relatively higher. The carapace sculpturing is similar to that of the male. The rostra are entire and lack the sulcus seen in males. They are also relatively longer (21.9 % CL) than in the male. The incisure is slightly shallower. The carapace glands are again large and open into a slot in the inner infold of the carapace 20 % of the carapace height below the posterior dorsal corner. The posterior dorsal corners form rounded right angles on both valves and there is a hint of a spine on the right valve.
Frontal organ. Absent
First antenna ( Fig. 7C, D View FIGURE 7 ). The limb is of much the same length as in the male, but the dorsal seta on the fifth segment is slightly longer, and the pad setae are slightly shorter. The terminal setae are also similar in length to those in the male.
Second antenna ( Fig. 7E, F View FIGURE 7 ). The protopodite is relatively shorter than in the male (31.4 % CL) as is the first segment of the exopodite (15.9 % CL). The first endopodite segment has a similar shape but is shorter than in the male (9.2 % CL). The longest setae on the endopodites on both limbs were broken, but the two shortest setae were shorter than in the male.
Mandible and maxilla. There is no sexual dimorphism in the structures of the mandible and maxilla. In the dissected female the terminal seta of the mandible was intact and was long (21.7 % CL) as illustrated by Scott (pl. IV, 7).
Fifth Limb ( Fig. 7G View FIGURE 7 ). The limb has three segments. The basale has a single dorsal plumose seta and a long, bare subterminal seta that extends to well beyond the end of the limb, but the suture just distal to their insertion is not shown in the figure. The first segment carries three lateral setae (2 + 1) and two pairs of ventral setae. The second segment has three ventral/lateral setae. The third segment has a pair of medial ventral setae and an adjacent lateral seta, and on the dorsal surface a short seta at two-thirds length. The terminal three setae are quite similar in length with the central longest seta being 11.5 % CL.
Sixth limb ( Fig. 7H View FIGURE 7 ). The limb also has three segments. The basale has the long bare dorsal seta that is a residual exopodite ( Boxshall 1998), and is subtended by a plumose seta. On its ventral margin the basale has three setae. The first endopodite segment has three pairs of ventral setae. The second segment has a pair of ventral setae at one-third length and a single dorsal seta at two-thirds length. Only two of the terminal setae were present on the sixth limb of the dissected specimen. The dorsal seta is not unusually long and is only slightly longer than the ventral seta.
Caudal furca ( Fig. 7I View FIGURE 7 ). There are eight pairs of claw setae, the longest of which is 13.4 % CL. Compared with other halocyprids, the claws do not diminish in length very much. The secondary spines are short and fine. There is a long unpaired seta.
Comparison between Bathyconchoecia paulula , S. darcythompsoni and S. arabica . Deevey (1968) designated Bathyconchoecia paulula as the type species for Bathyconchoecia . However, her description of B. paulula was based on specimens retrieved from the gut contents of benthic fish, so her descriptions are understandably often incomplete so that all her species need to be re-examined. Even so, comparisons between B. paulula and the two species described herein show they have many similarities, and so are closely related.
1. Offshore oceanic habitat.
2. The structure of the carapace with infolds and well defined sculpture.
3. The overall structure of the A1 is similar although the relative lengths of the setae may differ (direct comparisons are not possible as Deevey (1968) give no meristic data)
4. The rectangular shape and setation of the A2 endopodite and the shapes of the hook appendages. Deevey (1968) illustrates only two setae attached to the basal hasp of the right (fig. 1i) and left (fig. 1g), but the third setae had probably been lost.
5. The structure and setation of the mandibular limb (see her figure 2b), notably the three plumose setae that are residual features of the exopodite, the long subterminal dorsal seta on the first endopodite segment, the single ventral seta on the second segment, and the very long terminal seta on the third segment.
6. The structure and setation of the maxilla.
7. The setation of the fifth limb.
8. The structure of the caudal furca hook spines, which are slim, nearly straight and relatively evenly sized, and all have fine secondary spination.
However, there are also major differences between them, which are sufficient to classify them in different genera. These differences are summarised in Table 2. Four of these differences are particularly significant:
1. The structures of the toothed edges of the mandibular basale and coxale.
2. The structures of the rostra.
3. The positions on the carapace at which the carapace glands open.
4. The structures of the copulatory appendage.
Three other species currently classified as Bathyconchoecia species have similar characteristics to Scottoecia arabica , namely B. crosnieri Poulsen, 1969b , B. subrufa Angel, 1970 and B. foveolata Deevey, 1968 , and are classified in the new genus.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.