
Tamaricella cf. ribauti (Zachvatkin)
|
publication ID |
https://doi.org/10.5281/zenodo.203776 |
|
DOI |
https://doi.org/10.5281/zenodo.6193322 |
|
persistent identifier |
https://treatment.plazi.org/id/2D4B87A9-FFDC-F85F-FF2B-560AD4F3FEAB |
|
treatment provided by |
Plazi |
|
scientific name |
Tamaricella cf. ribauti (Zachvatkin) |
| status |
|
4) Tamaricella cf. ribauti (Zachvatkin) View in CoL
(Figs 10–12).
Material examined. CG: Sardegna (Cagliari): Stagno di Feraxi, 2,4 Km dalla SP 20; s.l.m.; 27/5/2006 (233); vegetation near lagoon with Inula , Phragmites, Salicornia , gramineae; Guglielmino & Bückle leg.; 26 males, 31 females.—Sardegna (Oristano): Mari Ermi; s.l.m.; 1/6/2006 (246); vegetation on seashore; Guglielmino & Bückle leg.; 23 males, 23 females.—Sardegna (Cagliari): Stagno di Feraxi, 2,4 Km dalla SP 20; s.l.m.; 31.03.2007 (338); vegetation near lagoon with Inula , Phragmites, Salicornia , gramineae; Guglielmino & Bückle leg.; 12 males; 9 females.—Sardegna (Oristano): Mari Ermi; s.l.m.; 11.04.2007 (359); vegetation on seashore; Guglielmino & Bückle leg.; 61 males, 50 females.—Lazio (Latina): Monte Circeo, c/o Terme Romane; 50–100m; 14/08/2001 (105); mixed forest near seashore with Crataegus , Pistacia lentiscus , Tamarix ; Guglielmino & Bückle leg.; 2 males, as T. tamaricis (Puton) .—Calabria (Cosenza): Trebisacce, Fiumara Fiume Saraceno near SS 106; ~ 20m; 16/ 06/2010 (537); dry areas with Pinus , Poaceae , Tamarix , Lamiaceae , Rosmarinus , Nerium oleander ; Guglielmino & Bückle leg.; 30 males, 45 females. CD: Sicilia (Catania): Misterbianco; on Tamarix ; 11/09/1987; D'Urso leg.; 1 male, as T. tamaricis (Puton) . CP: Sardegna (Oristano): Sinis, Turr'e Seu; 7.viii.1996; vegetation on seashore; F. Poggi leg.; 14 males, 17 females.—Sardegna (Oristano): Sinis, Is Arutas; 24.vii.1992; vegetation on seashore; F. Poggi leg.; 14 males, 12 females.
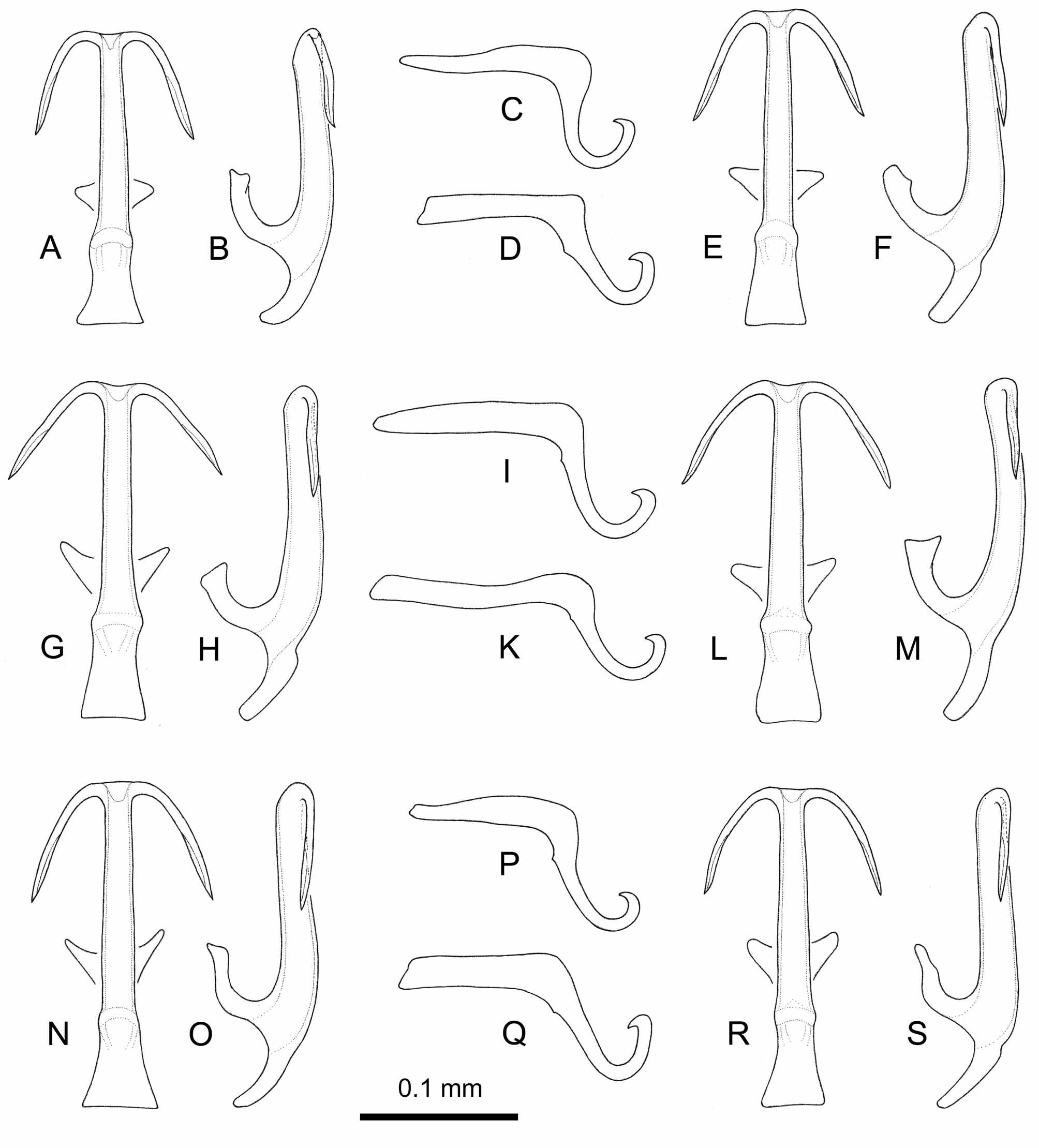
FIGURE 10. Tamaricella cf. ribauti (Zachvatkin) , male, Sardinia, Stagno di Feraxi: Aedeagus ventral (A); aedeagus lateral (B); right genital style from outside (C); apex of right genital style from outside (other specimen) (D); left pygofer process from outside (E); apical part of left pygofer process from outside (other specimen) (F); 2nd – 4th abdominal sterna, dorsal view (G).
(The locality numbers in parenthesis coincide with the number system used by Guglielmino and Bückle in their faunistic and zoogeographic papers).
Up to now in Italy only two Tamaricella taxa are recorded, both feeding on Tamarix : T. tamaricis (Puton) and a taxon identified as T. gr. cypria (Ribaut) . T. tamaricis is recorded from Basilicata, Calabria, Lazio, Sardinia, Sicilia (see Servadei 1967; D'Urso 1986; Guglielmino 1993; Guglielmino et al. 2005; Zocchi 1970). T. gr. cypria was found only in Pantelleria and Linosa (D’Urso & Guglielmino 1995).
Della Giustina (1989) records from Corsica T. remanei Dworakowska and figures details of the male genital apparatus and the apodemes of the 2nd abdominal sternum. His material was identified by Dworakowska, but there are slight differences, concerning above all the orientation of the apical aedeagal appendages, between the figures of Della Giustina (1989) and those given by Dworakowska (1971b) in her revision of the genus Tamaricella .
One of the authors (Poggi) in 1992 and 1996 in high summer collected specimens of a Tamaricella taxon in western Sardinia on the Sinis Peninsula on Frankenia laevis L.. During later collection trips Tamaricella populations were found by the two other authors in extremely high abundance at two places in Sardinia (one on the Sinis Peninsula) on Frankenia laevis L. as well, whereas no specimens of that genus seemed to be present on Tamarix bushes close to the Frankenia laevis localities. These results were obtained not only in spring but also in early summer. Frankenia laevis is a halophilous plant distributed near the seashore or lagoons around that island.
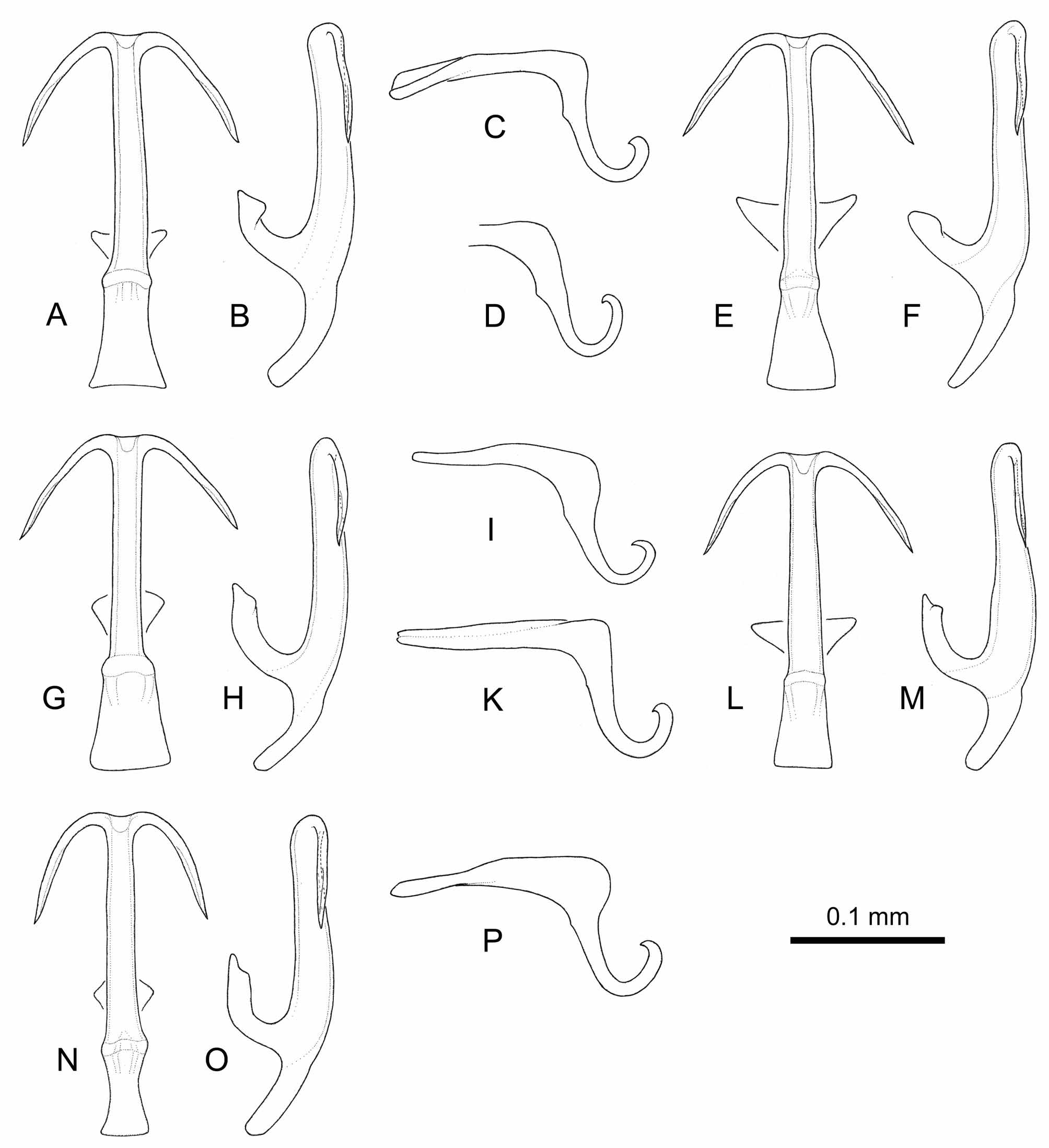
We compared the Sardinian Tamaricella specimens (Figs 10,11) collected on Frankenia laevis with specimens found on Tamarix spp. from the Italian mainland and Sicily: two males from M. Circeo (Lazio), recorded as T. tamaricis by Guglielmino et al. (2005) ( Figs 12 View FIGURE 12 N–P), one specimen from Misterbianco (Sicily), formerly identified as T. tamaricis as well ( Figs 12 View FIGURE 12 A–C), and a rich Tamaricella population from Torrente Saraceno (Calabria) ( Figs 12 View FIGURE 12 D–M).
Measurements (males). The specimens studied display a body length between 2.0– 2.65 mm. Thus, almost all specimens are clearly smaller than T. remanei (following Dworakowska (1971b) 2.65–2.85 mm) and more or less in the range of T. tamaricis or T. ribauti . Within the same population we observe often a distinctly reduced size range variability.
The measurements of the Sardinian specimens are interesting as they indicate different body size for spring and summer. The populations found in July and August on the Sinis Peninsula in Sardinia represented the smallest specimens with body size between 2.0 and 2.2 mm, head width between 0.61 and 0.65 mm ( 20 specimens measured). The largest specimens with body size between 2.35 and 2.65 mm, head width between 0.67 and 0.74 mm, were collected in spring ( March 31st and April 11th) in the same region ( Mari Ermi, 15 specimens measured) and at a locality in southeastern Sardinia (Feraxi, 9 specimens measured). At the last two places (at Feraxi) specimens were collected also at the end of May and the beginning of June. The measurements of these specimens (14 from Mari Ermi and 16 from Feraxi) resulted in body sizes mainly near those of the summer specimens from the Sinis Peninsula (body length 2.1–2.35 mm, head width 0.62–0.68 mm, with two specimens from Mari Ermi distinctly larger ( 2.5 mm / 0.70 mm and 2.55 mm / 0.72 mm). These two large specimens may represent late members of the spring generation, but it is not clear whether there is a definite sequence of generations or a continuous reproduction during the whole year.
Similar results as for the males were obtained also for the females.
The Calabrian population (on Tamarix sp., in June) contained specimens with body size between 2.3 and 2.5 mm, head width 0.66–0.69 mm ( 5 specimens measured), similar dimensions display the specimens from Sicily ( 2.5mm / 0.69 mm) and Latium ( 2.45–2.5 mm / 0.67–0.68 mm). Local and seasonal environmental factors may have influence on the body size. Thus, body size is perhaps not a very reliable diagnostic character.
Coloration. All the collected specimens display roughly the same coloration (in the specimens from Calabria the dark spots on pronotum are somewhat more marked): Head, pronotum and scutellum yellow with indistinctly delimitated olive green areas on disc of pronotum, vertex and sometimes frontoclypeus. Wings and legs olive green. Sterna and abdomen largely black. Markings: Dark markings below insertion of antennae, two rounded black spots on transition frontoclypeus-vertex surrounded by a light yellow band, four small dark spots behind middle half of anterior margin of pronotum, one indistinctly delimited sometimes almost lacking larger dark area on each side behind the eyes, often composed of several small spots, one of them linear comma-shaped and somewhat distant from anterior margin of pronotum, two triangular dark areas on scutellum with two very small spots between them.
Genital morphology (Figs 10–12). The aedeagus shows more or less variable features in the specimens we checked: the aedeagus shaft may be distinctly widened basally (lateral view, Fig. 10B) or evenly wide ( Fig. 11 View FIGURE 11 S), almost perfectly straight ( Fig. 11 View FIGURE 11 S) or distinctly curved ( Fig. 11 View FIGURE 11 M), the angle of shaft and its appendages in ventral view can vary as between Fig. 11 View FIGURE 11 A and Fig. 11 View FIGURE 11 G. In lateral view the appendages are constantly directed in parallel to the aedeagus shaft (as in Figs 10B, 11B, 12B); a very slight change of direction is perceptible at about midlength of the appendages and their apical part seems to be furrowed. Styles (Figs 10C, 10D) and apodemes of the 2nd sternum (Fig. 10G) are also somewhat variable. Pygofer appendages are to a conspicuous degree variable as well (Figs 10E, 10F), even if they display constantly a distally inwards bent tip. The variability in the above mentioned characters is observed also within the same population.
Morphologically all specimens seem to differ from T. tamaricis (see figures in Ribaut, 1936 and Dworakowska, 1977) by the distally inwards bent tip of pygofer appendages. Furthermore, they differ distinctly from the specimens of T. remanei figured by Della Giustina (1989) and Dworakowska (1977) in the orientation of the aedeagus appendages in lateral view. Even if single specimens share with T. remanei their basally enlarged aedeagus shaft, the character set consisting of body size, orientation of aedeagus appendages and shape of pygofer appendages, observed in the majority of the specimens collected in Italy, implies a close relationship to T. ribauti (Zachvatkin) and T. jaxartensis (Oshanin) .
Host plant. Our data from Sardinia imply that Frankenia laevis is the main host plant of the Tamaricella taxon collected there. The same plant genus is recorded as host plant at least for one other Tamaricella species, T. franckeniae Lindberg from the Canary Islands ( Lindberg 1953). In other places ( Portugal, Cape Verde Islands, see Lindberg 1958, 1960) Lindberg instead indicates the same species feeding on Tamarix . Frankeniaceae and Tamaricaceae are closely related families, which occur often in the same localities. A host plant shift from Frankenia to Tamarix or vice versa therefore would not be very surprising. Maybe our Sardinian taxon feeding on Frankenia , has the capacity of changing its host plant during the dry summer period with possible loss of, or at least unfavourable conditions on its main host plant. Vidano (1964) records T. tamaricis from three provinces of Sardinia and mentions only Tamarix spp. as host plant, he did not indicate the period during which his specimens were collected.
Remarks. In view of the slight differences between many Tamaricella taxa on the one hand and the conspicuous variability observed within the populations collected in Sardinia and in Calabria on the other, we consider necessary a revision of that species group defining clear diagnostic characters, based on large numbers of specimens of different populations with a detailed study of their range of variability and including data about their host plants. All records of T. tamaricis from Italy are to be controlled, in the present situation new records should contain in addition to a taxonomic attribution at least a sketch of pygofer appendages and aedeagus in lateral and ventral view. A definite identification of the Italian Tamaricella populations presently seems impossible. All the populations studied by the authors (except for T. gr. cypria from Pantelleria), collected in part on Frankenia laevis and in part on Tamarix spp. in Sardinia, Sicily and Italian mainland presently cannot be split up in different taxa. Provisionally we subsume them as T. cf. ribauti (Zachv.) . It is possible that there are two species, which we are unable to distinguish based on morphological characters, strictly associated with Tamarix sp. and Frankenia sp. respectively.
The high abundance often observed in Tamaricella populations, their well-developed flying ability, and their normal occurrence along rivers or seashore on Tamarix shrubs or on sandy open plains with Frankenia exposes them to air currents and storms which may easily transport swarming individuals to other sites suitable for colonization along valleys, the coastline, or to nearby islands; but this may not promote speciation. Different host plants could facilitate ecological isolation, which may explain the existence of several closely related taxa along the Mediterranean coasts, but our results in Italy do not support this hypothesis. Possibly some of the described taxa represent morphological variants of the same species and genetic data would help clarify the status of such taxa.
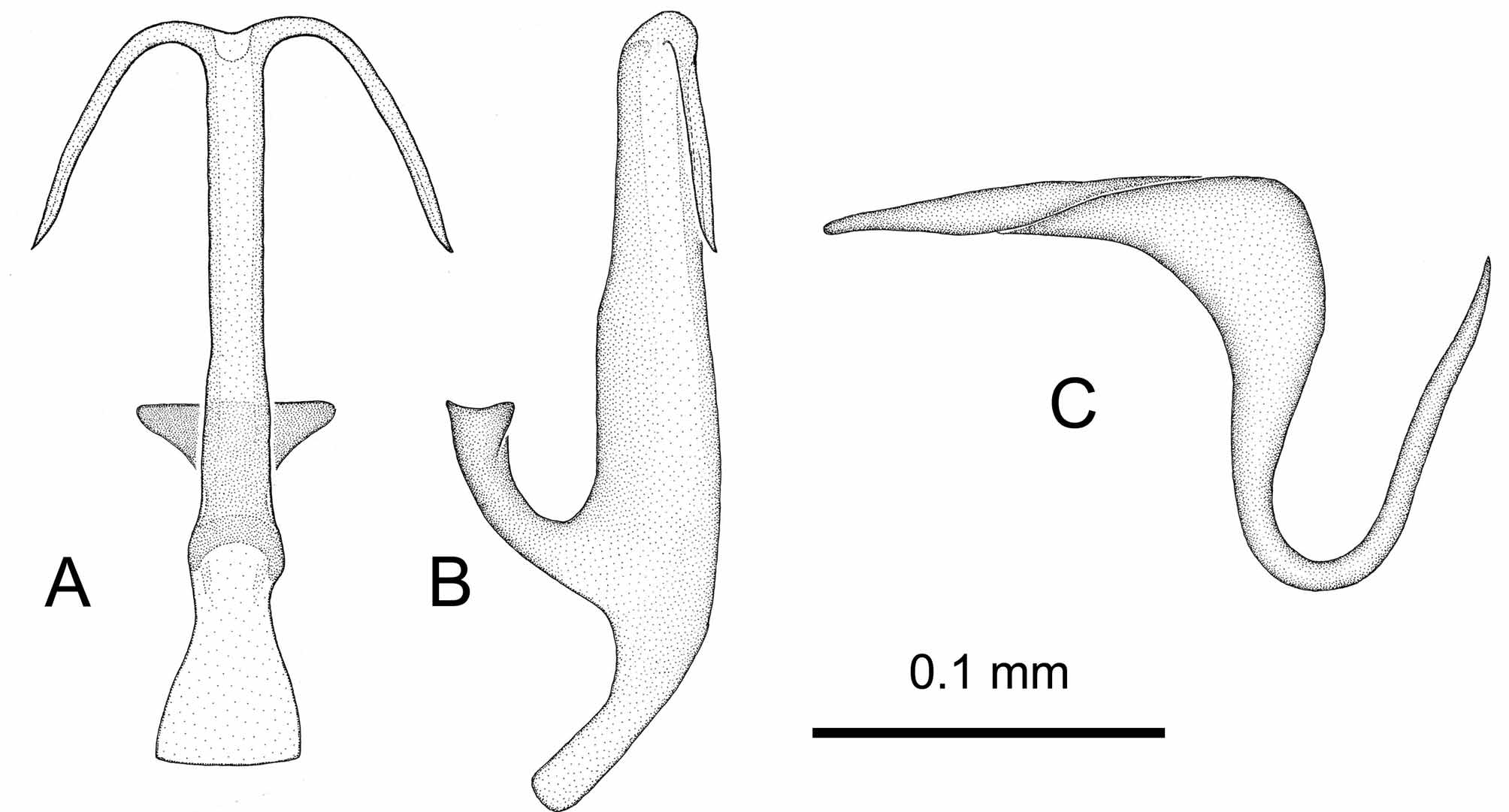
The single male specimen from Pantelleria (Mursia; on Tamarix ; 07.ix.1986; leg. A. Carapezza), deposited in Dipartimento di Biologia "Marcello La Greca", Università di Catania, Italy (collection D’Urso) (CD) and by D'Urso & Guglielmino (1995) recorded as " T. gr. cypria (Ribaut) " contrasts in its genital morphology markedly with all other Tamaricella specimens from Italy that we have examined ( Fig. 13 View FIGURE 13 ). The cypria -group comprises, apart from T. cypria (Ribaut) , recorded from Cyprus, Israel and Egypt, also T. franckeniae (Lindberg) , recorded from Canary Islands, Portugal and Cape Verde Islands, T. complicata Dworakowska recorded from Israel and Egypt, and T. lethierryi Dworakowska , recorded from Sudan and Eritrea. These species display, following the figures in Dworakowska (1971b), slight differences in the shape of the aedeagus shaft and of the pygofer appendages. The aedeagus shaft, which is abruptly thickened dorsally at midlength in lateral view ( Fig. 13 View FIGURE 13 B) and the long and slender, apically not curved pygofer appendages ( Fig. 13 View FIGURE 13 C) of our specimen from Pantelleria imply an attribution to T. complicata , but more material from Pantelleria should be studied to confirm this identification. Also in this case a revision of the species group, including studies on variability, food plant and genetic data would be desirable.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Typhlocybinae |
|
Genus |