
Natalus stramineus Gray, 1838
|
publication ID |
https://doi.org/ 10.1206/636.1 |
|
persistent identifier |
https://treatment.plazi.org/id/2C6DBE54-FFDA-FFE3-884C-FC06FE51B3CB |
|
treatment provided by |
Tatiana |
|
scientific name |
Natalus stramineus Gray, 1838 |
| status |
|
Natalus stramineus Gray, 1838 View in CoL Figure 33 View Fig
Natalus stramineus Gray, 1838: 496 View in CoL . Original description based on unspecified material of unknown geographic origin deposited at the BMNH. Handley and Gardner (1990) deduced that this taxon’s description must have been based on the specimen numbered BMNH 70.2324.
Natalus dominicensis Shamel, 1926: 67 . Subjective synonym. Original description based on material (USNM 113605) from the island of Dominica.
HOLOTYPE: Holotype by monotypy, BMNH 70.2324, adult male in alcohol with skull removed; collector and collection locality unknown. The skull lacks parts of the left temporal but is otherwise complete. The skin is in good condition although both tibiae and 15 finger bones are broken.
DISTRIBUTION: Islands of the Lesser Antilles north of the St. Lucia Channel: Anguilla, Antigua, Barbuda, Dominica, Guadeloupe, Marie Galante, Martinique, Montserrat, Nevis, Saba, and St. Maarten (skeletal remains only; fig. 23).
NOTE: The name Natalus stramineus has been historically applied to populations of the genus Natalus from virtually the entire Neotropics. The original description of Natalus stramineus ( Gray, 1838) , however, gives no indication of the geographic origin of the holotype, leading to widespread confusion concerning the nomenclature, and geographic and taxonomic limits of species of Natalus . The type locality of N. stramineus was listed as Brazil for about a century ( Cabrera, 1957; Carter and Dolan, 1978; Dobson, 1878). Despite claims that the type locality was in the Lesser Antilles ( Goodwin, 1959; Handley and Gardner, 1990), opinions that it could actually be in Brazil persisted ( Taddei and Uieda, 2001). This conflict was compounded by the fact that N. stramineus was assumed to range throughout South America, and to be sympatric with N. tumidirostris to the north of the Amazon ( Dávalos, 2005; Koopman, 1981; Simmons, 2005; Soriano and Ochoa, 1997). Recently, Tejedor (2006), on the basis of the inflated premaxilla of the holotype of N. stramineus , confirmed that the type locality of this species is in the northern Lesser Antilles (although the specific island remains uncertain) and showed that N. tumidirostris is the only taxon that occurs in South America north of the Amazon.
DIAGNOSIS: Medium-sized representative of genus Natalus (forearm length 37.2– 40.0 mm, breadth across canines (3.8– 4.2 mm) large in relationship to skull length; maxillary tooth row (6.8–7.5 mm) medial margin of ear deeply concave; lateral margin of ear deeply notched, 5–6 folds on lateral margin of ear; nostrils small, opening ventrolaterally; ventral hairs monocolored; dorsal hairs bicolored, hair bases lighter than tips, hair at base of claws short and inconspicuous or long and thin, never forming tufts; premaxilla inflated, with a lateral profile nearly forming a straight angle between rostral plane of premaxilla and dorsal plane of nasals; maxilloincisive suture dorsal to infraorbital foramen, maxilla dorsal to molars convex, not inflated; sides of postorbital region widely diverging rostrally, in dorsal view; palate between pterygoids ending caudally at two thirds of the distance between M3 and tip of pterygoid; posterior margins of maxilla, in ventral view, forming an oblique angle with longitudinal axis of skull; basisphenoid pits shallow; caudal margin of ascending ramus of mandible perpendicular to alveolar margin of dentary; I1 not visible in lateral view, obscured by I2; mesostylar crest on M3 absent. A comparison of diagnostic characters between N. stramineus , and other species of Natalus is summarized in table 5. View TABLE 5
DESCRIPTION: Medium-sized natalid (forearm length 36.9–41.9 mm; greatest skull length 15.7–17.5 mm; weight 4.6–5.3 g); muzzle long and dorsoventrally flattened; nostrils elliptical, opening ventrolaterally on shallow depression on margin of upper lip; upper lip thickened; lower lip markedly thickened and constricted along midline, with numerous transversal grooves; small, smooth central pad on dorsal margin of lower lip; natalid organ medium size and elliptical, extending from caudal base of rostrum to crown of head; ears relatively long (14.6– 17.5 mm); ear pinna funnel shaped but distally thin; pinna with markedly pointed tip; medial margin of pinna slightly to deeply concave; lateral margins of pinna deeply concave; five to six small ear ridges along lateral margin of distal pinna; ventral region of ear pinna greatly expanded, covering the eye and tragus in lateral view; medial ear margin thin and flexible; tragus short, lanceolate, and twisted into helixlike structure; tibia (19.8–23.2 mm) slightly longer than half the length of the forearm; calcar long and thin, occupying about half the length of the free edge of uropatagium; free margin of uropatagium with sparse fringe of thin hairs; wings relatively long and pointed, with 3rd metacarpal (33.8–38.1 mm) longer than 5th metacarpal (33.3–37.0 mm); wings attach to tibia above ankle; pelage dense, lax, and slightly woolly; hairs long (8–9 mm, dorsally; 6–7 mm, ventrally); dorsal hairs slightly bicolored, with bases lighter than tips; ventral hairs monocolored; dorsal hairs smoke gray with wood brown tips to amber or orange brown with medium brown tips (pl. 1); ventral hairs pale smoke gray or pinkish buff to light amber-chestnut or orange brown; dense mustachelike hair tufts along lateral margins of upper lip and on dorsum of muzzle; mustache formed by dense, lax, irregularly arranged, and ventrally curved hairs; natalid organ covered with thin hairs; skull long and relatively broad with moderate rostral flexion; rostrum wide and short, with sulcus between nasals shallow; moderate rostral palatal emargination; premaxilla moderately to markedly inflated; maxilloincisive suture slightly to markedly displaced caudally, sometimes dorsal to infraorbital foramen, in lateral view; maxilla convex above molars; braincase moderately inflated, rising gently from rostrum; sagittal crest moderately developed; postorbital constriction wide; maxillary branch of zygomatic arch thin, less deep than twice the height of crowns of last molars; pterygoids nearly parallel; palate extending caudally to more than half the distance between bases and tips of pterygoids; basisphenoid pit shallow; longitudinal medial ridge on basisphenoid present; ectotympanic small, covering less than half of periotic; upper incisors short and peglike; I2 obscuring I 1 in lateral view; occlusal profile of premolars long; upper premolars of similar size; mesostylar crests on M1 and M2 short and straight, mesostylar crest absent on M3; cingular cusp of p4 medium sized and broad; molars cusps relatively broad; spinous process of humerus about as high as capitulum; thorax relatively short and wide; ribs markedly expanded craniocaudally and extensively in contact with one another; vertebrae C7 to T1 fused among themselves and to ribs; vertebrae T12–L4 fused entirely without vestige of sutures; vertebrae L5 and L6 free; caudal vertebrae 4 to 7 longer than distance from ischium to iliac crest of sacrum.
COMPARISONS: The inflation of the premaxilla and the associated caudal displacement of the maxilloincisive suture is sufficient to distinguish N. stramineus from other species of Natalus as herein restricted. In most specimens of N. stramineus from the northern Lesser Antilles, the maxilloincisive suture is dorsal to the infraorbital foramen, whereas in specimens from the central Lesser Antilles and in all other species of Natalus this suture is located more anteriorly, at about 1/3 of the distance between the infraorbital foramen and the anteriormost margin of the premaxilla (plates 8, 14; table 1). Also, in specimens with an inflated premaxilla, in lateral view, the anterior and dorsal planes of the premaxilla form a nearly right angle in N. stramineus , whereas in all other Natalus this angle is obtuse, particularly so in N. tumidirostris . Specimens of N. stramineus that lack the inflated premaxilla can be distinguished from other species of Natalus by a combination of characters including size, discrete traits, and cranial proportions.
On the basis of body size alone, Natalus stramineus can be distinguished from the large Greater Antillean species N. primus, N. major , and N. jamaicensis . The observed range of forearm length in N. stramineus (37.2–41.9 mm) overlaps only slightly with that of N. major (41.1–45.0 mm, P, 0.01; table 1), the smallest of the Greater Antillean species. On the other hand, N. stramineus is slightly larger than the North American Natalus lanatus , and the North and Central American N. mexicanus (table 2), but from these species, N. stramineus is more easily distinguished by external and cranial characters. Whereas in N. stramineus the medial ear margin is concave (usually as much as the lateral ear margin), in N. lanatus it is straight. In addition, N. lanatus has shorter legs (length of tibia in fresh or fluid-preserved specimens, 16.2–19.2 mm) than N. stramineus (length of tibia 19.8–21.5 mm, P, 0.01). From N. mexicanus , N. stramineus can be distinguished by its upper incisors, which are at the same level in ventral view, whereas in N. mexicanus the caudal base of I1 is rostral to the caudal base of I2.
Natalus stramineus is most similar in size and external characters to the South American N. tumidirostris and N. espiritosantensis . From N. tumidirostris , N. stramineus can be separated by its small, oval, and ventrolaterally oriented nostrils (larger, usually circular, and more anteriorly facing in N. tumidirostris ) and by a palate extended caudally between the pterygoids (the palate is emarginated in N. tumidirostris ). Relative to N. stramineus , N. espiritosantensis has a more robust, shorter rostrum (reflected in its shorter maxillary tooth row, P, 0.01; table 5 View TABLE 5 ).
VARIATION: The sample of Natalus stramineus examined for this study showed significant sexual, geographic, and individual variation. Males were significantly larger than females in two external and four cranial dimensions (MANOVA, P, 0.05; table 20). Canonical variate analyses between four geographic samples ( Anguilla, Antigua + Barbuda, Saba + Montserrat, Dominica), using four external and seven cranial dimensions, indicate that the sample from Dominica is morphometrically distinct from that of the remaining islands, in particular with longer 3rd metacarpals ( fig. 34A View Fig ) and narrower breadths across canines ( fig. 34B View Fig ). The three available skins from Anguilla are grayish brown (smoke gray at base and wood brown at tips, with an overall drab appearance), whereas those from the remainder of the range have a reddish amber brown to tawny olive brown pelage.
The morphology of the rostrum showed both geographic and individual variation. The inflation of the premaxilla that characterizes the holotype of N. stramineus was present in all specimens from Anguilla, Barbuda, and Antigua, and in one female from Saba and another from Montserrat. Two adult females and one adult male from Saba, however, lacked this trait, being indistinguishable, at least cranially, from specimens from Guadeloupe and Dominica.
NATURAL HISTORY AND CONSERVATION: Natalus stramineus is known from 35 localities (including three represented only by bone remains), 15 of which have been caves. It is found both in the volcanic innerarc islands (Saba, Nevis, Montserrat, Dominica, and Martinique) and in the limestone outer-arc islands ( Anguilla, St. Maarten, Barbuda, Antigua, Guadeloupe (Grande-Terre), Marie Galante; Baker et al., 1978; Genoways et al., 2001; Pedersen et al., 2003, in press). In Dominica, in addition, it has been taken in a brick tunnel ( Genoways et al., 2001). On Nevis, it has been found in a small, humid cave (3 m wide by 6 m high) that is not permanently occupied throughout the year ( Pedersen et al., 2003). In Dominica, it has been found coexisting with several thousand Pteronotus davyi in a narrow cave opening on a sandy cliff ( Genoways et al., 2001).
Natalus stramineus occurs from sea level to middle elevations and in habitats ranging from relatively dry (e.g., Barbuda, 925 mm annual precipitation; locality 7) to rain forest (e.g., Sylvania, Dominica, 3232 mm; locality 171) and from sea level to 473 m (locality 171).
Although its diet has never been studied, N. stramineus is surely insectivorous, as are other representatives of Natalidae (e.g., Nyctiellus lepidus, Silva-Taboada, 1979 ; Natalus primus, Tejedor et al., 2004 ).
It is known from 114 museum specimens, most of which are from the islands of Dominica (77), Barbuda (12), and Saba (8), where it seems to be a common bat. The records from Marie Galante, and Martinique, the last of which is known from a mention in A. Schwartz’s research notes ( Timm and Genoways, 2003), are based on a single specimen each.
Little is known of reproductive activity in N. stramineus but it may be concentrated earlier in the year than in other species of Natalus from the West Indies. Females taken in April in Dominica included lactating, nonpregnant, and pregnant individuals, yet a sample from July was composed entirely of nonreproductive females (Genoways et al.,
2001). N. stramineus is parasitized by the streblid Trichobius caecus ( Genoways et al., 2001) .
In an ecomorphological study, Jennings et al. (2004) stated that N. stramineus stramineus can fly very slowly in clutter and that it hunts by slow hawking and/or by gleaning. They found that N. stramineus produced multiharmonic FM echolocation calls with most energy in the second harmonic. The
greatest energy of the fundamental frequency was centered around 60 kHz, and that of the second harmonic at 90–100 kHz. The duty cycle was intermediate ( Jennings et al., 2004).
Natalus stramineus View in CoL is listed as least concern in the current global red list of threatened species (IUCN, 2010) However, its small, naturally fragmented geographic range, and its apparent requirement for humid caves, point to a limited availability of suitable habitat and therefore to a vulnerable status. The species seems, nonetheless, locally common in at least four islands ( Antigua, Barbuda, Dominica and Saba; Genoways et al., 2001; S.C. Pedersen, personal commun.), and future surveys may increase the number of islands where it is known to occur. On Nevis, it has been encountered less frequently, with the single specimen in museum collections from that island having been killed accidentally by a ceiling fan ( Pedersen et al., 2003). In Martinique, it is also known from a single individual ( Timm and Genoways, 2003), and in St. Maarten, it has been recorded exclusively from a skull and partial skeleton found in a cave ( Genoways et al., 2007). In Montserrat, an island where bat populations have been severely affected by recent volcanic eruptions (Pedersen, 2003), the only known population of N. stramineus View in CoL occurs in Aymers Ghaut (S.C. Pedersen, in litt.), one of the island’s valleys most vulnerable to devastating pyroclastic flows and lahars ( Baker, 1985). If N. stramineus View in CoL does not occur in other, undiscovered sites on Montserrat, its long-term survival on that island seems highly uncertain.

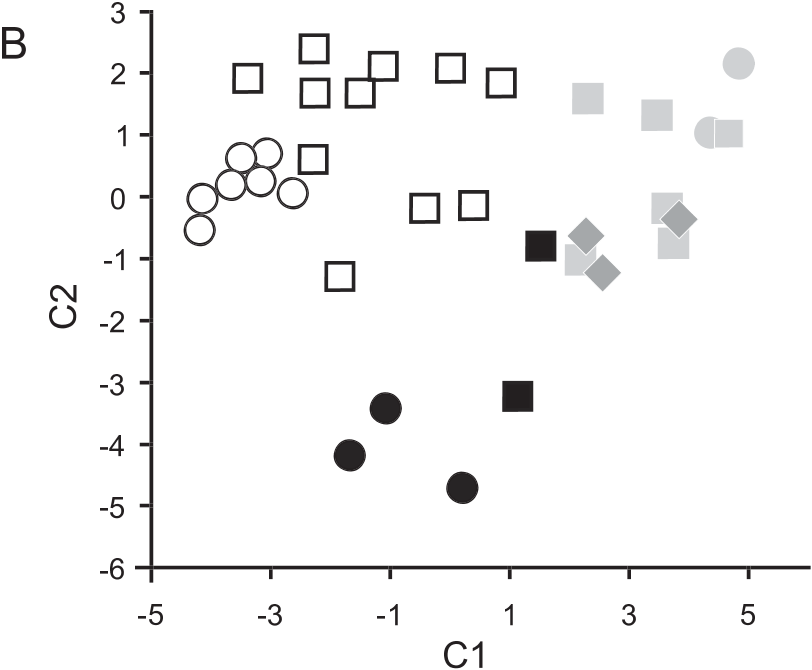
TABLE 20 Summary of measurementsa of Natalus stramineus
| Females | Males | |||||||
|---|---|---|---|---|---|---|---|---|
| N | Mean | (Min.–Max.) | SD | N | Mean | (Min.–Max.) | SD | |
| Weight | 4 | 4.9 | (4.6–5.3) | 0.3 | 3 | 5.0 | (4.9–5.2) | 0.2 |
| Forearm length | 26 | 38.9 | (36.9–41.2) | 1.1 | 39 | 39.5 | (37.8–41.9) | 0.9 |
| Length of tibia, dry* | 6 | 19.2 | (18.8–19.8) | 0.4 | 8 | 20.1 | (18.3–21.7) | 1.1 |
| Length of tibia* | 21 | 20.9 | (19.8–22.0) | 0.8 | 23 | 21.7 | (20.8–23.2) | 0.6 |
| Length of 3rd metacarpal | 18 | 36.5 | (33.8–38.1) | 1.1 | 20 | 36.9 | (35.5–37.9) | 0.8 |
| Length of 5th metacarpal* | 18 | 35.2 | (33.3–36.7) | 0.8 | 20 | 35.9 | (35.1–37.0) | 0.5 |
| Length of ear | 12 | 16.0 | (14.6–17.5) | 0.7 | 15 | 16.0 | (14.7–17.5) | 0.8 |
| Length of penis | – | – – | – | 20 | 3.2 | (2.4–4.2) | 0.5 | |
| Length of natalid organ | – | – – | – | 14 | 6.7 | (5.2–7.8) | 0.9 | |
| Greatest skull length | 13 | 16.3 | (15.7–17.5) | 0.5 | 24 | 16.7 | (15.9–17.2) | 0.3 |
| Zygomatic breadth* | 14 | 8.3 | (7.9–8.8) | 0.2 | 24 | 8.5 | (7.9–8.9) | 0.2 |
| Braincase breadth | 13 | 7.8 | (7.6–8.4) | 0.2 | 23 | 8.0 | (7.8–8.3) | 0.1 |
| Breadth across molars* | 14 | 5.4 | (5.1–5.6) | 0.2 | 24 | 5.6 | (5.0–5.9) | 0.2 |
| Breadth across canines* | 14 | 3.7 | (3.5–4.1) | 0.2 | 23 | 3.9 | (3.3–4.2) | 0.2 |
| Maxillary tooth row | 14 | 7.0 | (6.8–7.5) | 0.2 | 24 | 7.3 | (6.9–7.5) | 0.2 |
| Mandibular tooth row | 14 | 7.5 | (7.3–7.8) | 0.2 | 17 | 7.6 | (7.2–7.8) | 0.1 |
| Postorbital breadth | 14 | 3.1 | (3.0–3.3) | 0.1 | 24 | 3.2 | (3.0–3.4) | 0.1 |
| Depth of braincase* | 8 | 6.4 | (6.2–6.7) | 0.2 | 17 | 6.6 | (6.4–6.8) | 0.1 |
a Descriptive statistics of measurements for each sex. N 5 sample size; SD 5 standard deviation. Weight is given in g; all other measurements are given in mm. See text for description of measurement methods. Measurements significantly different between sexes (P, 0.05) are followed by an asterisk (*).
TABLE 21 Loadings of the first two canonical axes of two canonical variates analyses (CVA) performed with 4 geographic samples of Natalus stramineus
| Measurement | C I | CII |
|---|---|---|
| External | ||
| Forearm length | 21.646 | 0.227 |
| Length of tibia | 0.382 | 20.881 |
| Length of 3rd metacarpal | 1.825 | 0.524 |
| Length of 5th metacarpal | 20.289 | 20.762 |
| Eigenvalues | 3.997 | 0.598 |
| Cranial | ||
| Greatest skull length | 20.340 | 0.228 |
| Zygomatic breadth | 20.806 | 0.447 |
| Braincase breadth | 0.748 | 0.477 |
| Breadth across molars | 20.197 | 0.485 |
| Breadth across canines | 1.101 | 21.339 |
| Maxillary tooth row | 0.886 | 0.723 |
| Postorbital width | 20.670 | 20.149 |
| Eigenvalues | 8.938 | 2.976 |
CVA scores are plotted in figure 49.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.